Acid Protease Enzyme for Effective Ethanol Fermentationは、穀物・植物性原料中のタンパク質をペプチドやアミノ酸へ低分子化し、酵母が利用しやすい窒素環境を整えるための酸性プロテアーゼです。エタノールを直接生成する酵素ではなく、糖化酵素や酵母の働きを栄養面・原料分解面から補助する工程用酵素として位置づけられます。穀物系発酵では、プロテアーゼを含む酵素処理が原料ポリマーの変換や酵母代謝に関与し得ることが報告されています[1]。
エタノール発酵の主役は、発酵性糖をエタノールと二酸化炭素へ変換する酵母です。デンプン系原料であれば、アミラーゼやグルコアミラーゼがデンプンを糖へ変換し、セルロース系原料であればセルラーゼ、ヘミセルラーゼ、β-グルコシダーゼなどが多糖を分解します。酸性プロテアーゼの役割はこれらとは異なり、原料に含まれるタンパク質画分を加水分解して、ペプチド、遊離アミノ酸、その他の可溶性窒素成分を増やすことにあります[2]。
穀物原料、豆類副産物、食品残渣、リグノセルロース系バイオマスには、炭水化物だけでなくタンパク質、フィチン酸、ミネラル結合成分、細胞壁マトリックスが共存しています。糖化だけを最適化しても、酵母が利用できる窒素源が不足すれば、発酵初期の増殖、糖消費速度、ストレス耐性、副生成物プロファイルに影響が出ます。酸性プロテアーゼは、こうした「糖はあるが酵母栄養が十分でない」状態を緩和するために使われます[3]。
Enzymes.bioは、酸性プロテアーゼを含むプロテアーゼ製品をオンライン供給する事業者であり、製造業者や研究所としてではなく、B2B用途の酵素供給元として製品を提供します。製品は1kg単位でオンライン直接購入でき、注文時にはCoAとSDSが併せて提供されます。特定工程での性能は原料、酵母株、pH、温度、保持時間、併用酵素に依存するため、本稿では過度な収率保証ではなく、酸性プロテアーゼがエタノール発酵で果たす実際的な機能に焦点を当てます。
トウモロコシ、小麦、ライ麦、ソルガムなどの穀物発酵では、デンプンが主要な炭素源ですが、タンパク質画分も発酵性能に関わります。穀物タンパク質は加熱、粉砕、糖化だけで完全に酵母利用可能な形になるとは限りません。酸性プロテアーゼは、貯蔵タンパク質や可溶性タンパク質のペプチド結合を切断し、酵母が取り込みやすい低分子窒素成分を増やす方向に働きます[1]。
酵母は糖を消費するだけでなく、細胞増殖、膜成分合成、酵素合成、酸化ストレス応答のために窒素を必要とします。アミノ酸や短鎖ペプチドが適切に供給されると、発酵初期の細胞数形成や代謝の立ち上がりを支えやすくなります。一方で、窒素源が不均一な場合、糖が残っていても発酵速度が低下したり、酵母がストレス応答へ傾いたりすることがあります[3]。
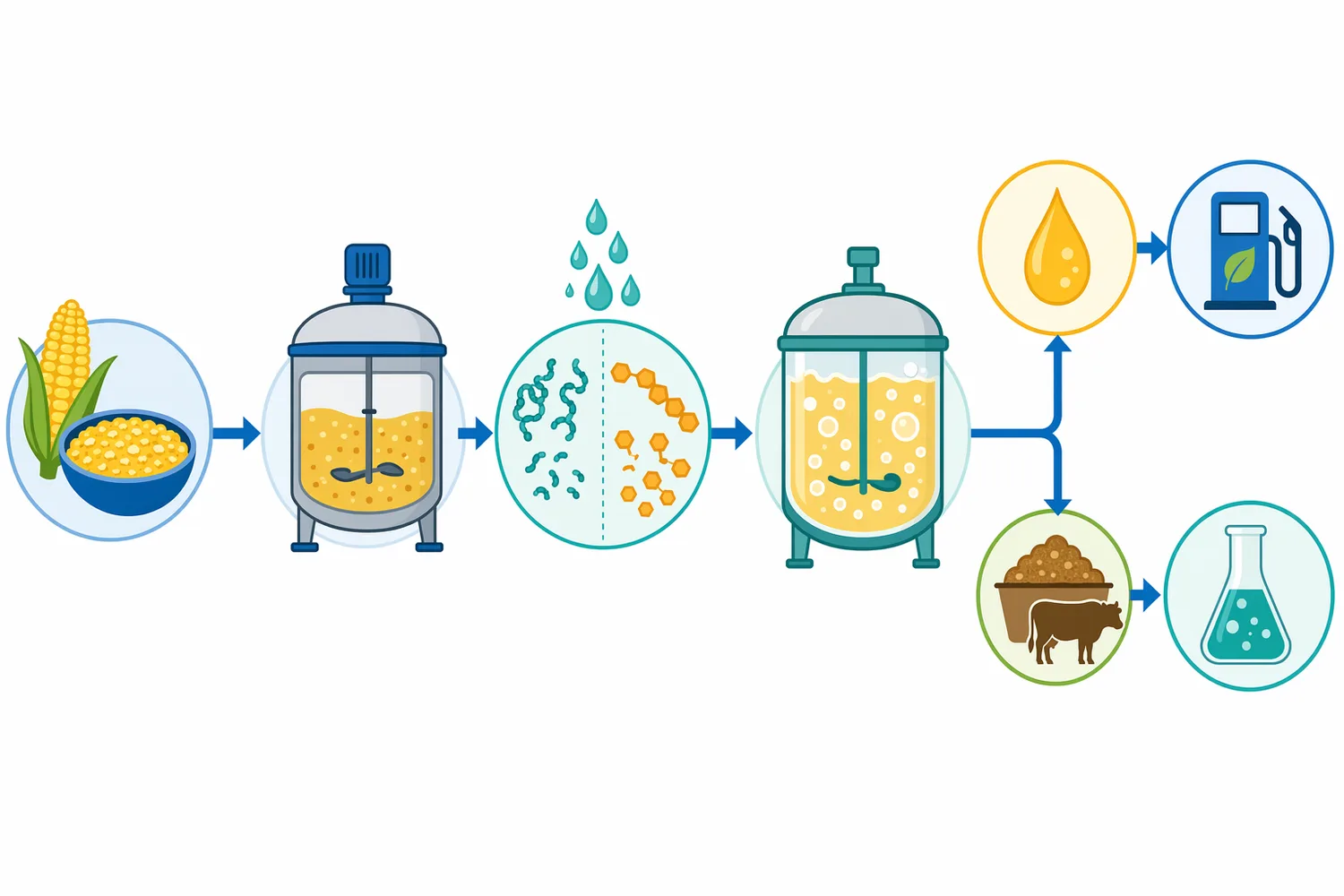
高固形分発酵では、原料濃度を上げることで設備当たりのエタノール生産性を高められる可能性がありますが、同時にスラリー粘度、混合、熱移動、酵素拡散、酵母への阻害が問題になりやすくなります。タンパク質がデンプン粒や細胞壁成分と複合体を作っている場合、糖化酵素が基質にアクセスしにくくなることもあります。酸性プロテアーゼによるタンパク質低分子化は、原料構造を緩め、他の加水分解酵素が作用しやすい環境を作る補助要因になります[4]。
リグノセルロース系原料では、前処理、酵素糖化、発酵を組み合わせる設計が一般的です。Miscanthus x giganteusを対象とした研究では、穏和条件での前処理、酵素加水分解、エタノール発酵が一連の工程として検討されています。酸性プロテアーゼはセルロースやヘミセルロースを糖へ変える酵素ではありませんが、複合バイオマス中のタンパク質画分を処理することで、総合的な原料分解戦略の一部になり得ます[4]。
エタノール濃度が上がる発酵後半では、酵母は膜流動性の変化、酸化ストレス、浸透圧、栄養枯渇など複数の負荷を受けます。酸性プロテアーゼはエタノール耐性を直接付与する酵素ではありません。しかし、発酵初期から中期にかけて利用しやすい窒素源を形成することで、酵母が十分な細胞機能を整えた状態で高エタノール環境へ進むことを補助します[5]。
特に、外部栄養剤だけに頼らず原料由来タンパク質を有効活用したい工程では、酸性プロテアーゼの意味が明確になります。タンパク質を未分解の固形分として残すより、ペプチドやアミノ酸として発酵液へ移行させる方が、酵母にとって利用可能な栄養プールが広がります。これは、発酵の安定性を「酵母株だけ」でなく「原料側の可利用性」から設計する考え方です[1]。
プロテアーゼは、タンパク質中のペプチド結合を加水分解する酵素です。エンド型プロテアーゼはタンパク質鎖の内部を切断し、タンパク質を中〜短鎖ペプチドへ変換します。エキソ型ペプチダーゼは末端側からアミノ酸を放出し、低分子化をさらに進めます。酸性プロテアーゼは、酸性寄りの環境でこうした反応を担う酵素群であり、発酵前処理や酸性マッシュのような条件で機能しやすいことが特徴です[6]。
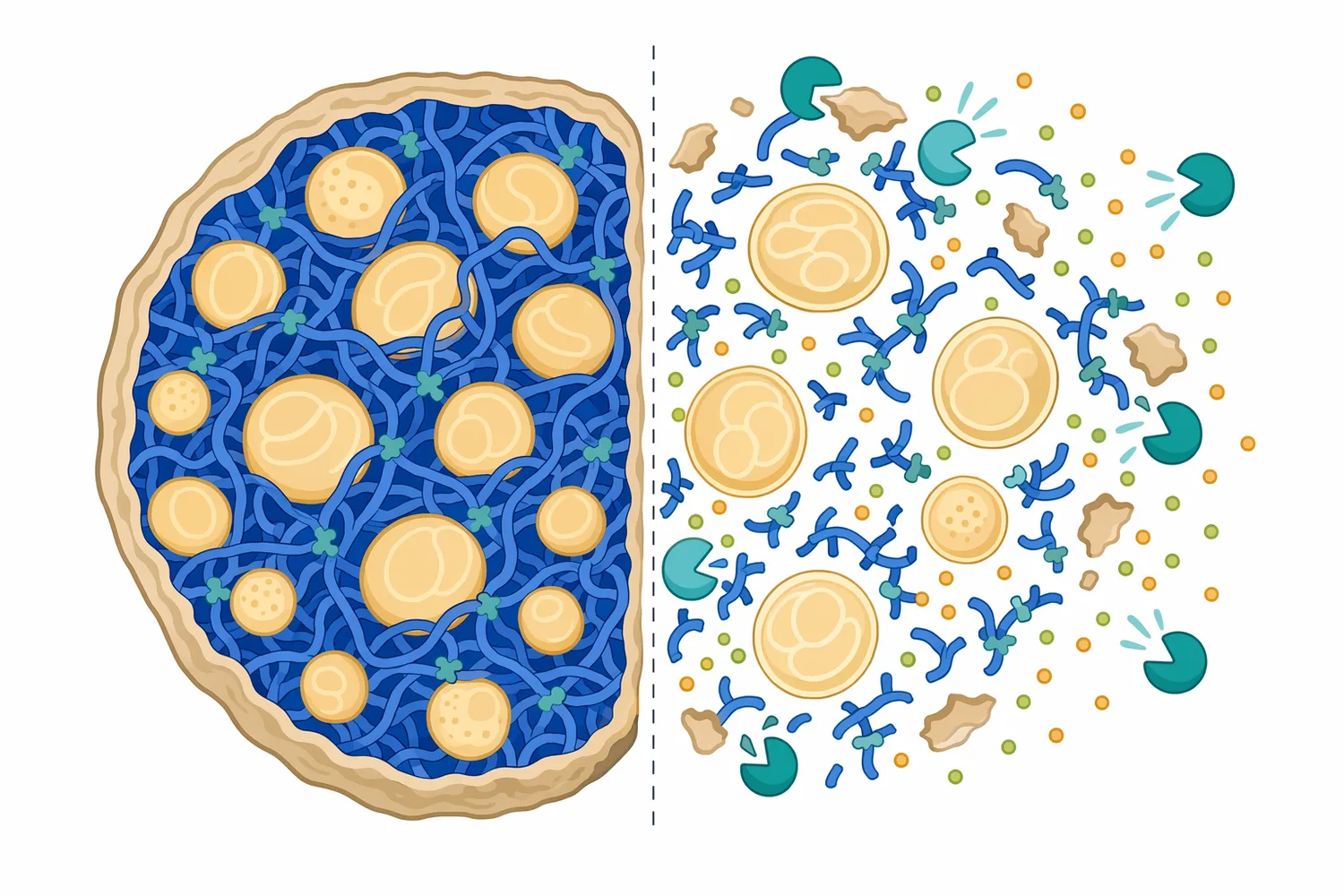
エタノール発酵の観点では、タンパク質分解そのものが最終目的ではありません。重要なのは、分解によって生成するペプチドやアミノ酸が、酵母の窒素代謝に接続されることです。酵母はアミノ酸を取り込み、タンパク質合成、酵素合成、細胞分裂、レドックスバランスの調整に利用します。また一部のアミノ酸は、発酵副生成物や香気成分の前駆体にもなり得ます[3]。
穀物や植物性原料では、タンパク質は単独で存在するのではなく、デンプン粒、細胞壁、多糖、脂質、ミネラルとともに複雑なマトリックスを形成しています。酸性プロテアーゼがこのタンパク質部分を切断すると、原料粒子の表面や内部構造が変化し、糖化酵素がデンプンや多糖へ接近しやすくなる場合があります。この効果は、酸性プロテアーゼが糖を生成するという意味ではなく、他の加水分解反応を妨げる物理的・化学的要因を緩めるという意味です[7]。
トリチケールわらの研究では、アルカリ前処理、酸加水分解、酵素加水分解、エタノール発酵が比較・最適化の対象になっています。こうした研究は、発酵収率が単一酵素だけで決まるのではなく、前処理、糖化、発酵の組み合わせによって左右されることを示しています。酸性プロテアーゼも同様に、単独で工程を完結させる酵素ではなく、多酵素設計の中でタンパク質画分を担当する要素です[7]。
Saccharomyces cerevisiaeなどの酵母は、糖を解糖系でピルビン酸へ変換し、低酸素条件ではピルビン酸脱炭酸酵素とアルコール脱水素酵素を介してエタノールを生成します。この炭素フラックス自体は酸性プロテアーゼが直接触媒するものではありません。しかし、酵母細胞がこの代謝を継続するには、酵素タンパク質、膜輸送体、ストレス応答タンパク質を維持する必要があり、その材料として窒素源が必要です[5]。
アミノ酸供給は、発酵の副生成物にも影響します。ワイン発酵のレビューでは、Saccharomyces cerevisiaeが揮発性化合物を形成する生化学的・分子機構が整理されており、アミノ酸代謝が発酵香気に関与することが示されています。燃料用エタノールでは香気は主目的ではありませんが、この知見は「アミノ酸は単なる栄養素ではなく、酵母代謝全体に関与する基質である」ことを理解する助けになります[3]。
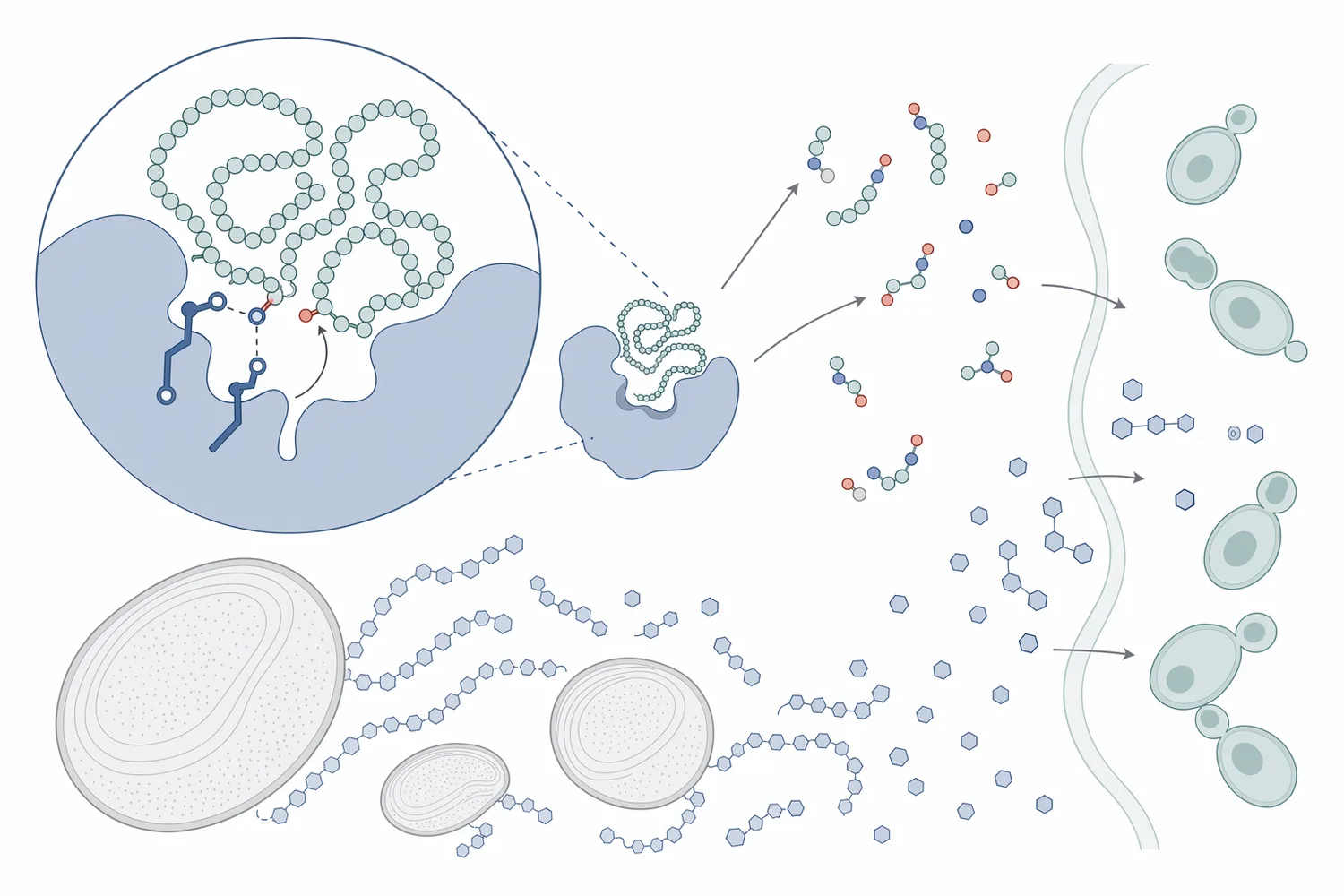
酸性プロテアーゼは、糖化酵素や繊維分解酵素と混同されがちですが、基質も生成物も異なります。エタノール発酵工程では、各酵素がどの原料画分を処理するのかを分けて考えることが重要です。
| 酵素の種類 | 主な基質 | 主な生成物 | エタノール発酵での役割 | 酸性プロテアーゼとの関係 |
|---|---|---|---|---|
| 酸性プロテアーゼ | タンパク質、ペプチド | ペプチド、アミノ酸、可溶性窒素 | 酵母栄養、原料マトリックス緩和 | 窒素利用性を補助する中心酵素 |
| α-アミラーゼ | デンプン | デキストリン、短鎖糖 | 液化・デンプン分解 | 炭素源生成を担当し、役割が異なる |
| グルコアミラーゼ | デキストリン、オリゴ糖 | グルコース | 発酵性糖の供給 | 酵母への糖供給を担当 |
| セルラーゼ系 | セルロース | セロビオース、グルコース | セルロース系原料の糖化 | 複合バイオマスで併用される |
| ヘミセルラーゼ系 | キシランなど | キシロース、オリゴ糖 | 副糖の生成、構造緩和 | 植物細胞壁分解で補完関係 |
| フィターゼ | フィチン酸 | リン酸、ミネラル可利用化成分 | ミネラル・栄養利用性の改善 | 穀物原料でプロテアーゼと組み合わせられることがある |
この比較から分かるように、酸性プロテアーゼは「糖を作る酵素」ではありません。エタノール収率に直結する発酵性糖は、主にアミラーゼ、グルコアミラーゼ、セルラーゼ、ヘミセルラーゼによって供給されます。酸性プロテアーゼは、酵母がその糖を効率的に利用するための栄養環境を整え、原料中のタンパク質を有効な発酵資源へ近づけます[8]。
酵素使用量は、バイオ燃料工程の経済性にも関係します。デアセチル化とディスクリファイニング工程の技術経済分析では、精製エネルギーや酵素使用が糖およびエタノールの販売価格指標に影響する要因として扱われています。これは、酵素を多く入れればよいという単純な話ではなく、どの酵素がどのボトルネックを解くのかを工程全体で考える必要があることを示しています[8]。
デンプン系原料では、エタノール発酵の中心はデンプン液化、糖化、酵母発酵です。酸性プロテアーゼはこの主経路に対して、タンパク質分解という補助経路を提供します。たとえば小麦やトウモロコシでは、タンパク質がデンプン粒や細胞壁成分と結びつくことで、糖化効率やマッシュ性状に影響する場合があります。酸性プロテアーゼは、こうしたタンパク質を低分子化し、可溶性窒素を増やす目的で利用されます[1]。
醸造分野では、麦芽や副原料中のタンパク質分解が遊離アミノ態窒素、発酵性、泡、濁りなどに関わることが知られています。燃料用エタノールでは官能品質は主目的ではありませんが、酵母が利用する窒素成分を原料から作るという考え方は共通しています。酸性プロテアーゼは、低pH寄りのマッシュ条件でタンパク質画分を処理しやすい点で、穀物発酵に組み込みやすい酵素です[1]。
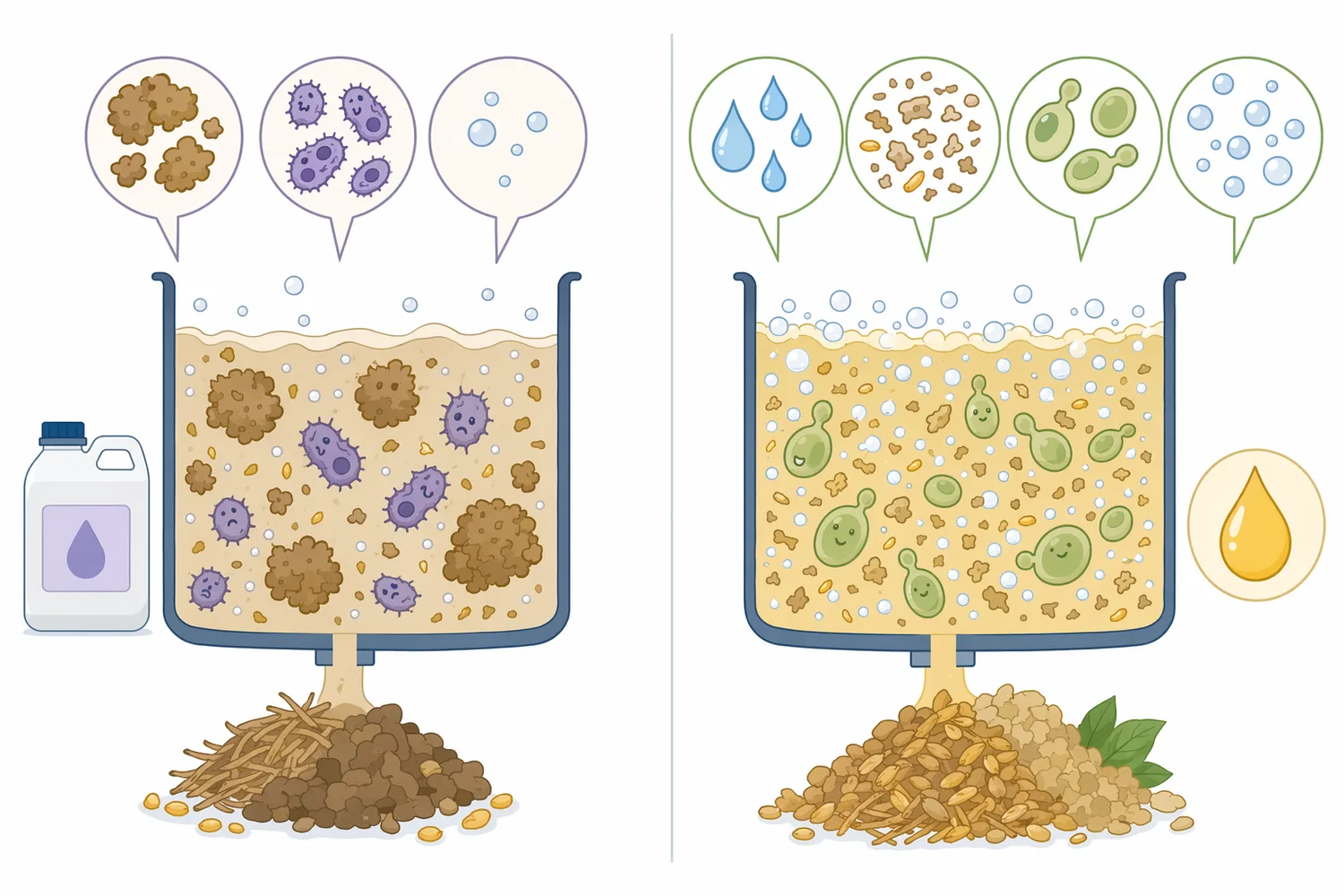
ソルガムやトリチケールのような原料では、デンプン、可溶糖、繊維、タンパク質の比率が品種や処理条件で変わります。甘味ソルガムを対象とした研究では、酸触媒加水分解と同時糖化発酵を組み合わせてエタノール生産を高める条件が検討されています。こうした原料では、酸性条件、糖化、発酵が密接に関係するため、酸性域で働くタンパク質分解酵素は補助酵素として検討しやすい位置にあります[9]。
トリチケールわらの研究では、前処理条件と酸・酵素加水分解の組み合わせがエタノール発酵に接続されています。わらや茎葉系原料は炭水化物だけでなく構造性成分が多く、単一酵素で完全に分解することは困難です。酸性プロテアーゼは、セルラーゼやヘミセルラーゼの代替ではなく、複合原料中のタンパク質部分を担当する補助機能として整理するのが妥当です[7]。
リグノセルロース系バイオマスでは、セルロース、ヘミセルロース、リグニンの相互作用が糖化を制限します。前処理によって構造を開き、酵素加水分解で糖を放出し、その後に酵母または改変酵母が発酵します。酸性プロテアーゼはリグニンを分解する酵素ではなく、セルロースを直接糖化する酵素でもありませんが、タンパク質や酵素阻害性タンパク質様成分が関与する原料では、補助的に意味を持つ可能性があります[4]。
キシロース利用性酵母の構築とエタノール発酵に関する研究は、リグノセルロース原料ではグルコース以外の糖利用が重要になることを示しています。酸性プロテアーゼはキシロース代謝を改善する酵素ではありませんが、酵母が複数糖を利用する高負荷発酵では、窒素環境とストレス耐性の確保がより重要になります[5]。
酸性プロテアーゼについて確実に言えるのは、タンパク質を加水分解し、ペプチドやアミノ酸を生成する酵素であるという点です。酸性プロテアーゼを生産・最適化・特性評価した研究では、糸状菌由来酵素が固体発酵によって得られ、酸性プロテアーゼとして評価されています。これは、酸性プロテアーゼが発酵産業で利用される酵素群として研究対象になっていることを示します[6]。
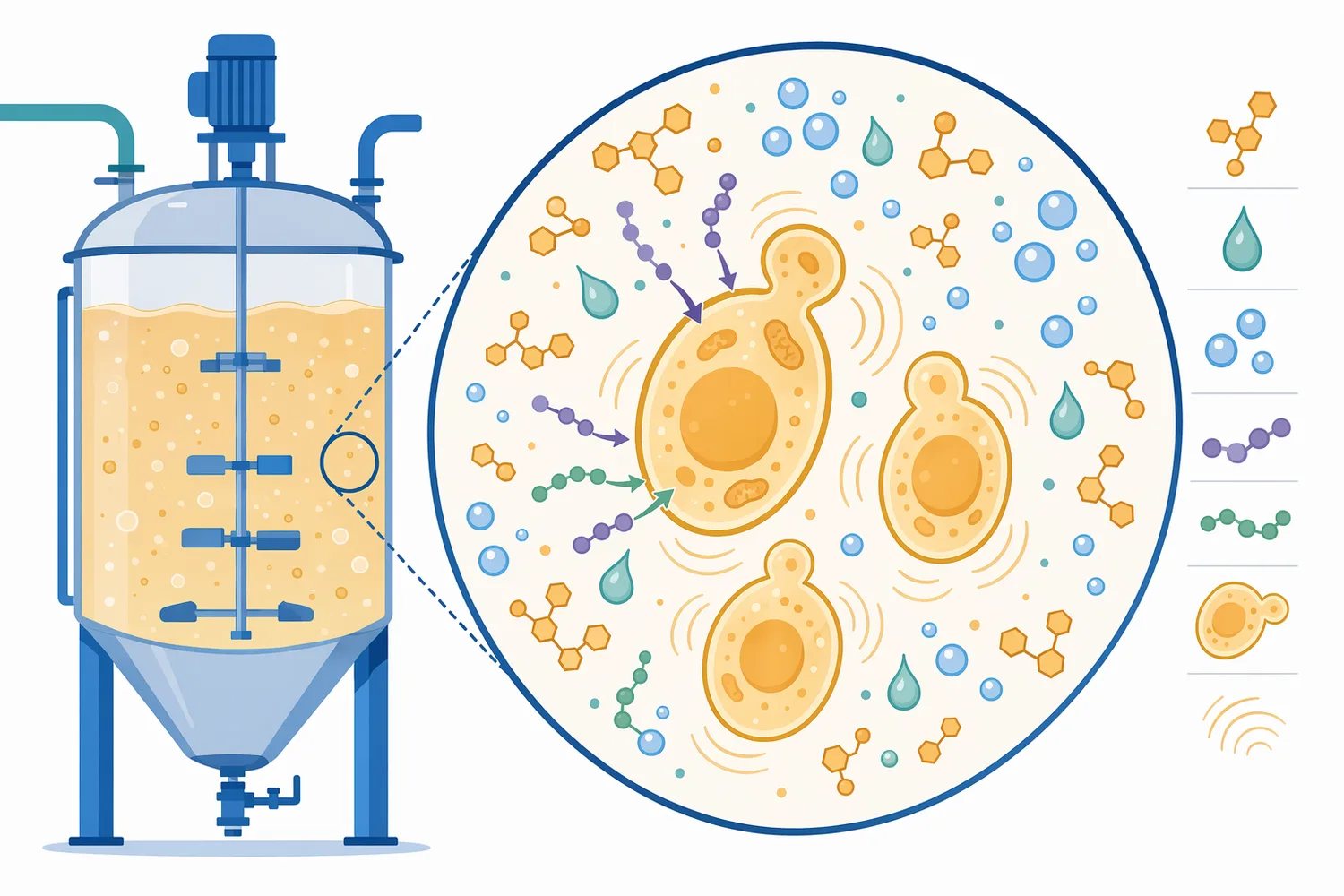
一方で、「酸性プロテアーゼを添加すれば、あらゆる原料でエタノール収率が一定割合上がる」とは言えません。エタノール収率は、糖化可能な炭水化物量、前処理、酵母株、阻害物質、発酵温度、pH、固形分濃度、栄養状態に左右されます。酸性プロテアーゼは、このうち主にタンパク質分解と窒素利用性に関与する補助因子であり、糖不足や酵母阻害を単独で解決するものではありません[8]。
また、プロテアーゼ処理によってアミノ酸が増えれば常に望ましいとは限りません。アミノ酸組成や濃度は酵母代謝、副生成物、発酵液の化学バランスに影響します。ワイン発酵の研究領域では、酵母のアミノ酸代謝が揮発性化合物形成に関与することが整理されています。燃料用途では香味は問題になりにくいものの、発酵代謝が窒素源の質に反応するという点は同じです[3]。
酸性プロテアーゼは、原料が水和され、タンパク質が酵素に接触できる状態で機能します。したがって、スラリー化後、糖化前、糖化中、または発酵前処理の段階で、タンパク質分解の時間を確保する設計が一般的に考えられます。発酵後半はエタノール濃度が上がり、酵母と酵素の双方にとって環境が厳しくなるため、酸性プロテアーゼの主な効果は発酵の初期設計に置く方が合理的です[5]。
同時糖化発酵では、糖化酵素と酵母が同じ系内で働くため、酸性プロテアーゼも併用酵素の一部として扱われます。ただし、酸性プロテアーゼは糖化酵素の代わりではないため、デンプン系原料ではアミラーゼ系、セルロース系原料ではセルラーゼ系との役割分担が必要です。甘味ソルガムの研究では、酸触媒加水分解と同時糖化発酵の最適化がエタノール生産と結び付けて検討されており、工程統合の重要性を示しています[9]。
酸性プロテアーゼを工程に組み込む際の基本は、原料タンパク質を「発酵阻害にもなり得る未利用固形分」ではなく、「酵母が利用可能な窒素資源」へ近づけることです。この考え方は、糖化収率だけでなく、酵母の生育、発酵速度、残糖、発酵完了性を総合的に見る工程設計と相性があります[1]。
第一の利点は、原料由来窒素の利用性向上です。タンパク質を低分子化することで、酵母が取り込みやすいペプチドやアミノ酸を増やし、発酵初期の栄養状態を整えます。これは、外部から窒素栄養を補う考え方とは異なり、原料中にすでに存在するタンパク質をより使いやすい形へ変換するアプローチです[2]。
第二の利点は、多酵素処理における補完性です。アミラーゼ、グルコアミラーゼ、セルラーゼ、ヘミセルラーゼが炭水化物画分を処理するのに対し、酸性プロテアーゼはタンパク質画分を担当します。リグノセルロース系原料や副産物原料では、単一の酵素機能ではなく複数の加水分解機能を組み合わせることが重要であり、酸性プロテアーゼはその一部として機能します[4]。
第三の利点は、原料ロット差への対応力です。穀物や農産副産物では、タンパク質含量、粒度、熱履歴、乾燥条件、貯蔵状態がロットごとに変わります。酸性プロテアーゼによってタンパク質分解の工程を持たせると、原料中の窒素成分を可溶化する方向に働くため、酵母栄養のばらつきを緩和する設計がしやすくなります[1]。
第四の利点は、発酵工程の経済性を考える際の選択肢になることです。酵素使用はコスト要因ですが、適切な酵素が糖化や発酵のボトルネックを緩和すれば、工程全体の価値に影響します。技術経済分析では、酵素使用量が糖やエタノールの価格指標に関係する要素として扱われており、酸性プロテアーゼも「どの課題に対して使うか」を明確にすることが重要です[8]。
Enzymes.bioは、プロテアーゼ酵素をオンラインで提供する供給業者です。酸性プロテアーゼは、エタノール発酵において原料タンパク質の分解と酵母栄養の改善を目的として使用される工程補助酵素として位置づけられます。製品は1kg単位でオンライン直接購入でき、注文時にCoAとSDSが併せて提供されます。
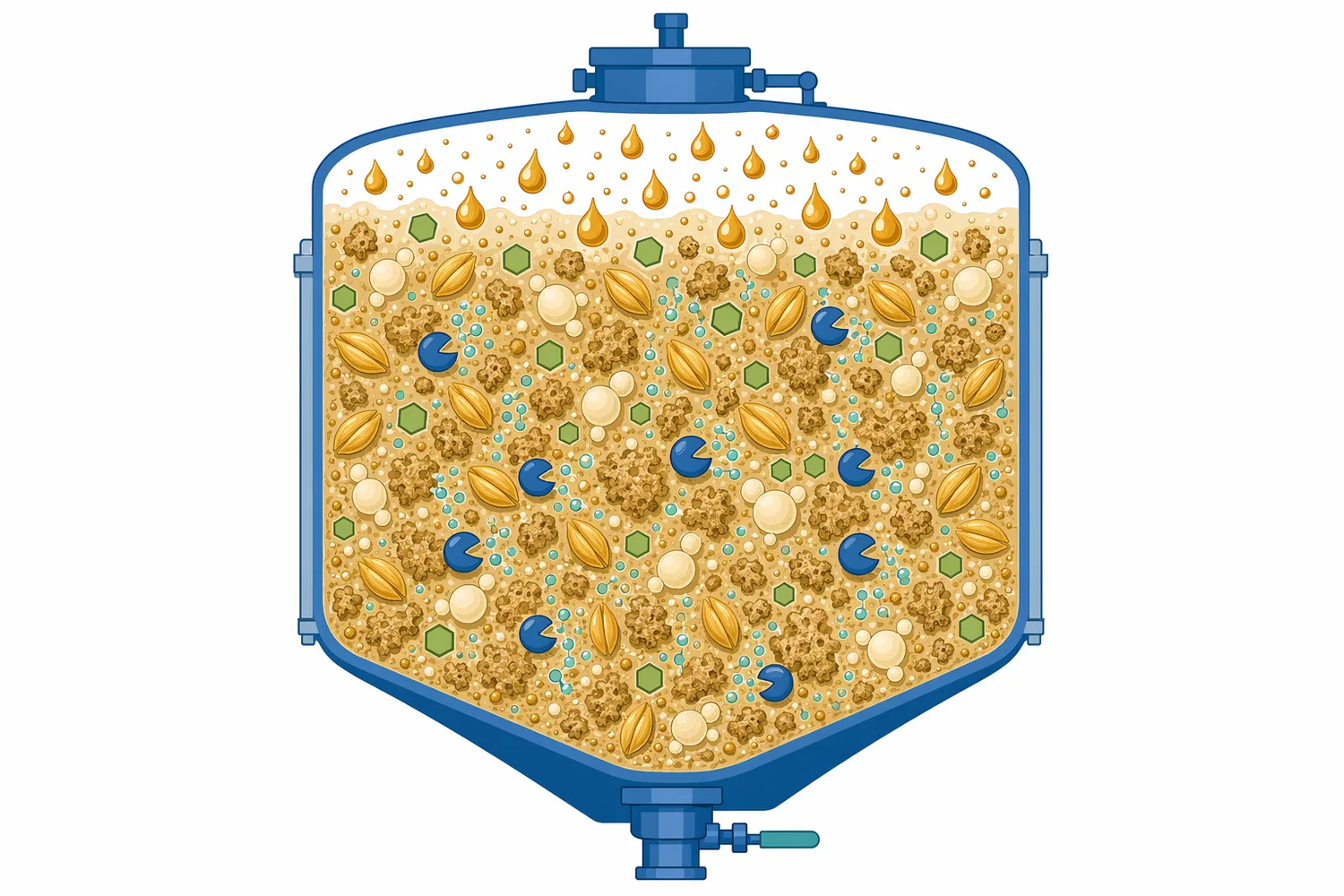
本稿は、購入前後の技術理解を助けるための解説であり、Enzymes.bioが製造試験や研究評価を代行することを示すものではありません。また、特定の活性単位、分析法、グレード、工程性能をここで規定するものでもありません。酸性プロテアーゼの価値は、発酵性糖の直接生成ではなく、タンパク質分解を通じて酵母が働きやすい原料環境を形成する点にあります[6]。
Acid Protease Enzyme for Effective Ethanol Fermentationは、エタノール発酵工程でタンパク質画分を低分子化し、ペプチドやアミノ酸を増やすための酸性プロテアーゼです。糖化酵素や酵母の代替ではなく、酵母が発酵性糖を安定して利用するための窒素環境を整える補助酵素として理解するのが正確です[2]。
穀物、ソルガム、トリチケール、リグノセルロース系原料、副産物系原料では、炭水化物の糖化だけでなく、タンパク質や細胞マトリックスの扱いが発酵挙動に影響します。酸性プロテアーゼは、アミラーゼ、グルコアミラーゼ、セルラーゼ、ヘミセルラーゼ、フィターゼなどと役割を分けながら、原料の総合的な加水分解に寄与します[9]。
最も実務的な表現をすれば、酸性プロテアーゼは「エタノールを直接増やす添加剤」ではなく、「原料タンパク質を酵母が利用しやすい窒素資源へ変換し、発酵工程を支える酵素」です。Enzymes.bioでは、この酸性プロテアーゼを1kg単位でオンライン供給し、注文時にCoAとSDSが提供されるため、B2Bの発酵用途で必要な基本文書とともに導入できます。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Acid Protease Enzyme For Effective Ethanol Fermentationを購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。