Alpha-Galactosidase(α-ガラクトシダーゼ)は、ラフィノース、スタキオース、ガラクトマンナン側鎖などに含まれるα結合型ガラクトース残基を加水分解する酵素です。大豆、豆類、植物性タンパク質、飼料原料、糖蜜、ガム質多糖の工程で、発酵性、粘度、消化時の不快感、基質利用性に関わる糖質組成を調整する目的で利用されます[1]。Enzymes.bioは製造業者や研究所ではなく、B2B向け酵素供給業者としてAlpha-Galactosidaseを1kg単位でオンライン販売し、CoAとSDSは注文時に併せて提供されます。
α-ガラクトシダーゼの実務上の価値は、「植物性原料に残りやすいα-ガラクトシド系糖質を、より単純な糖または低分子糖質へ変換すること」にあります。豆類に含まれるラフィノースファミリーオリゴ糖(RFOs)は、種子の成熟・発芽でα-ガラクトシダーゼ活性と連動して変動することが示されており、α-ガラクトシダーゼが植物性糖質の動態に直接関わる酵素であることが分かります[2]。
一般検索では「alpha galactosidase」「alpha galactosidase beano」「alpha galactosidase lipase alpha amylase protease and lactase capsules uses」「alpha galactosidase cofactors」などが見られますが、B2Bの食品・飼料・工業加工では、サプリメント的な説明よりも、基質、結合様式、工程条件、原料マトリックスとの相互作用を明確にすることが重要です。経口α-ガラクトシダーゼについては、Beanoのような消費者向け製品を対象にした二重盲検クロスオーバー試験もありますが、これは工業用酵素原料を直接摂取用として説明する根拠にはなりません[3]。
α-ガラクトシドは、ガラクトースがα配置で他の糖や糖鎖に結合している構造を指します。豆類や大豆原料で重要になる代表例はラフィノースファミリーオリゴ糖で、ラフィノース、スタキオース、ベルバスコースなどはヒトや一部動物の消化管で処理されにくく、後段の発酵やガス発生に関係することがあります。エンドウ種子の研究では、ラフィノースオリゴ糖の分解阻害が発芽遅延につながることが示され、RFO分解が生理的にも機能的にも重要な反応であることが確認されています[1]。
α-ガラクトシダーゼは、これらの糖質に含まれるα-D-ガラクトース残基を認識し、加水分解によってガラクトースを遊離させます。ササゲ種子を対象にしたα-ガラクトシド加水分解の速度論的研究では、豆類中のα-ガラクトシドが酵素反応で分解される対象として扱われており、食品・植物原料加工での基質理解に近い文脈を提供しています[4]。
α-ガラクトシダーゼは、デンプン、タンパク質、脂質を広く分解する酵素ではありません。α-アミラーゼがデンプンのα-グルカン結合を、プロテアーゼがペプチド結合を、リパーゼが脂質エステル結合を、ラクターゼすなわちβ-ガラクトシダーゼが乳糖のβ-ガラクトシド結合を主対象にするのに対し、α-ガラクトシダーゼはα結合型ガラクトースを含む糖質を主に標的とします。β-ガラクトシダーゼは乳糖不耐や乳糖加水分解の文脈でレビューされており、α-ガラクトシダーゼとは基質選択性が異なります[5]。
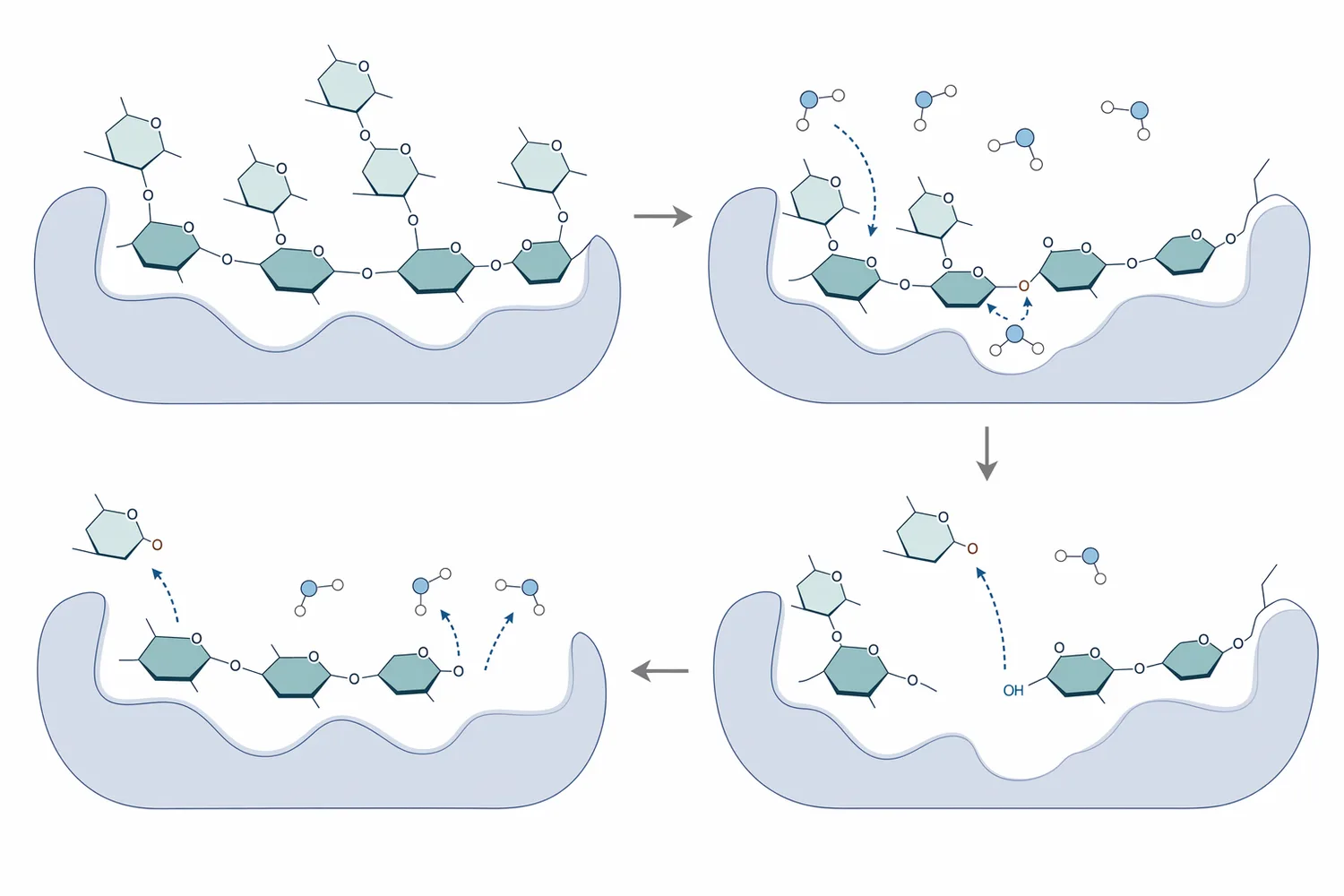
この違いは、「alpha galactosidase lipase alpha amylase protease and lactase capsules uses」のような複合酵素検索語を読む際にも重要です。複数酵素を並べた製品名では一括りに見えますが、工業工程では各酵素が触媒する結合、作用する原料画分、望ましい反応タイミングが異なります。β-ガラクトシダーゼは食品、医療、工業で多用途に利用されますが、乳糖やガラクトオリゴ糖に関する反応が中心であり、豆類RFOやガラクトマンナン側鎖の処理を説明するにはα-ガラクトシダーゼの基質特異性を分けて理解する必要があります[6]。
大豆、ヒヨコマメ、エンドウ、ササゲなどの豆類原料では、ラフィノースファミリーオリゴ糖が加工品質と消化時の印象に影響します。ヒヨコマメでは、種子成熟と発芽の過程でα-ガラクトシダーゼ活性が空間的に制御され、RFO量に影響することが示されています。これは、豆類原料中のRFOが単なる副成分ではなく、酵素活性と連動して変化する糖質群であることを示す実験的根拠です[2]。
食品工程でのα-ガラクトシダーゼ利用は、RFOを低減して原料を扱いやすくする方向に設計されます。たとえば豆乳、植物性飲料、豆ペースト、植物性タンパク質分散液では、原料中の可溶性糖質が風味、発酵挙動、後味、腸内発酵に関わるため、α-ガラクトシダーゼ処理は糖質組成を狙って調整する手段になります。経口用途で「ガスを防ぐか」を評価したBeano関連研究は消費者製品の文脈ですが、α-ガラクトシダーゼが食事性オリゴ糖不耐の課題と結びつけて研究されてきたことを示しています[3]。
植物性タンパク質原料では、タンパク質濃縮だけでなく、随伴する糖質や繊維画分が製品物性に影響します。α-ガラクトシダーゼ処理は、タンパク質そのものを分解するのではなく、共存するα-ガラクトシド系糖質を変換するため、プロテアーゼ処理とは目的が異なります。食品廃棄物や複合原料の酵素加水分解に関するレビューでは、pH、温度、基質濃度、酵素と基質の接触性が加水分解効率に影響する要因として整理されており、植物性原料でも同様にマトリックス条件が重要です[7]。
代替肉、植物性飲料、発酵豆乳、植物性プロテイン粉末では、糖質組成のわずかな違いが風味や微生物発酵の立ち上がりに影響します。α-ガラクトシダーゼは、強酸や過度な熱処理で糖質を非選択的に分解するのではなく、特定のガラクトシド結合に作用するため、原料ダメージを抑えたい食品工程に適しています。ただし、最終効果は原料の前処理、加水状態、熱履歴、糖質の溶出度、混合条件によって変動します[7]。
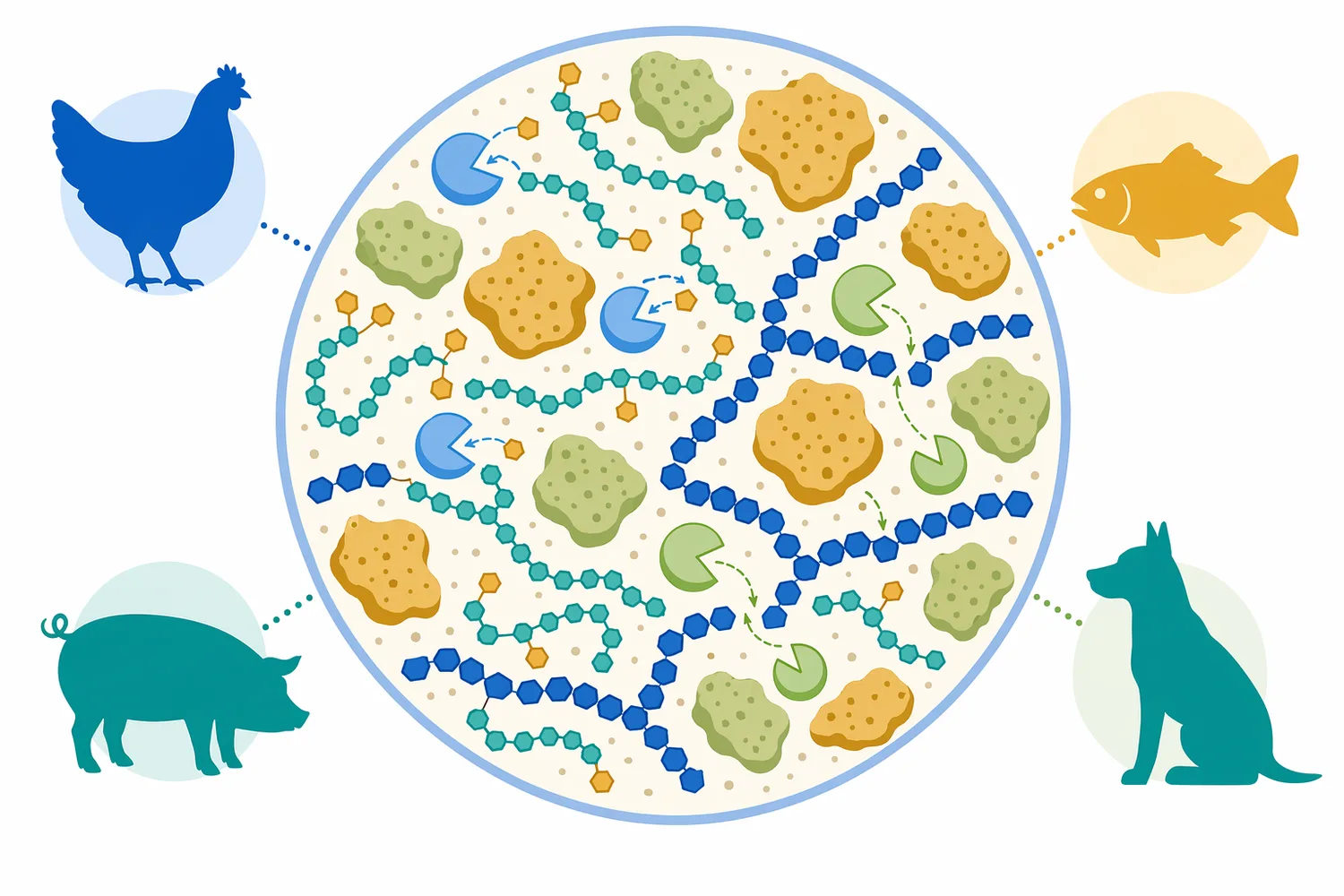
飼料分野では、大豆粕、豆類、植物性副産物などの利用効率が重要です。α-ガラクトシダーゼは、動物が利用しにくいα-ガラクトシド系糖質を低分子化する補助酵素として検討されます。マンナン分解に関するレビューでは、β-マンナナーゼ、β-マンノシダーゼ、α-ガラクトシダーゼの相乗作用が取り上げられており、α-ガラクトシダーゼがガラクトマンナン側鎖の除去を通じて主鎖分解を助ける役割を担うことが示されています[8]。
この相乗性は、単一酵素だけで複雑な植物細胞壁や貯蔵多糖を完全に処理しようとするより、基質構造に応じて複数の糖質分解酵素を組み合わせる考え方に通じます。ガラクトマンナンでは、マンナン主鎖に付いたガラクトース側鎖が他の酵素のアクセスを妨げることがあり、α-ガラクトシダーゼが側鎖を外すことで、β-マンナナーゼによる主鎖切断が進みやすくなります[8]。
α-ガラクトシダーゼは、豆類RFOだけでなく、ガラクトマンナン系多糖の処理にも関係します。Thermotoga neapolitana由来の高耐熱性α-1,6-ガラクトシダーゼとβ-1,4-マンナナーゼは、グアーガム加水分解用の酵素ブレーカーとして研究されており、高温環境でのガラクトマンナン処理にα-ガラクトシダーゼが関与し得ることを示しています[9]。
グアーガラクトマンナン溶液をガラクトマンナナーゼとα-ガラクトシダーゼの混合系で処理した研究では、加水分解中のレオロジー特性が評価されています。粘度や流動性は食品、飼料、発酵、油田関連など多様な工程で操作対象になるため、α-ガラクトシダーゼは単なる「オリゴ糖分解酵素」ではなく、多糖構造の側鎖処理を通じて物性に影響し得る酵素としても位置づけられます[10]。
ビート糖蜜をα-ガラクトシダーゼとエタノール生産へ変換する研究では、糖蜜のような糖質豊富な副産物を微生物・酵素プロセスに接続する文脈でα-ガラクトシダーゼが扱われています。糖蜜には発酵に利用しやすい糖と利用しにくい糖が混在するため、α-ガラクトシドを変換する酵素は、発酵基質の可用性を変える要素になります[11]。
この用途では、α-ガラクトシダーゼが直接エタノールを作るのではなく、微生物が利用しやすい糖質組成へ基質を整える前処理または併用酵素として機能します。発酵工程では、糖の種類、濃度、浸透圧、阻害成分、微生物種が結果を左右するため、α-ガラクトシダーゼの役割は「発酵の万能促進剤」ではなく、「特定糖質の変換によって発酵可能性を変える酵素」と表現するのが正確です[11]。
| 用途領域 | 主な基質・原料 | α-ガラクトシダーゼが担う反応 | 工程上の狙い | 根拠の読み方 |
|---|---|---|---|---|
| 大豆・豆類加工 | ラフィノース、スタキオースなどのRFO | α結合型ガラクトース残基の加水分解 | ガス発生要因、発酵性、糖質組成の調整 | 種子・豆類研究でRFO分解とα-ガラクトシダーゼ活性の関係が示される[1] |
| 植物性タンパク質 | タンパク質画分に随伴する可溶性糖質 | 糖質画分の選択的変換 | 風味、後味、消化時印象、発酵挙動の調整 | 効果はマトリックス、pH、温度、接触性に依存する[7] |
| 飼料・動物栄養 | 大豆粕、豆類、ガラクトマンナン含有原料 | 難利用性糖質や側鎖ガラクトースの処理 | 原料利用性、腸内発酵挙動の調整 | マンナン分解酵素群との相乗作用が報告される[8] |
| グアーガム・ガラクトマンナン | ガラクトース側鎖を持つマンナン | α-1,6ガラクトース側鎖の除去 | 粘度、流動性、主鎖分解の補助 | 高耐熱酵素ブレーカーやレオロジー研究がある[9] |
| 糖蜜・発酵基質 | 糖蜜中の複合糖質 | 発酵可能糖への変換補助 | 微生物利用性の改善 | ビート糖蜜の生物変換研究で扱われる[11] |
α-ガラクトシダーゼは「ガラクトースに関係する酵素」という点でβ-ガラクトシダーゼと混同されやすいですが、両者は結合様式が異なります。β-ガラクトシダーゼは乳糖加水分解やガラクトオリゴ糖生成の中心酵素であり、固定化β-ガラクトシダーゼの食品産業応用も広くレビューされています[12]。
| 酵素 | 主な標的結合・基質 | 代表的な用途 | α-ガラクトシダーゼとの違い |
|---|---|---|---|
| α-ガラクトシダーゼ | α-D-ガラクトシド、RFO、ガラクトマンナン側鎖 | 豆類糖質低減、飼料、ガム質多糖処理 | α結合型ガラクトースを標的にする |
| β-ガラクトシダーゼ/ラクターゼ | 乳糖などのβ-D-ガラクトシド | 乳糖分解、GOS生成 | 乳糖処理が中心で、α-ガラクトシド処理とは別機能[5] |
| α-アミラーゼ | デンプンのα-グルカン結合 | 糖化、製パン、醸造、澱粉加工 | ガラクトース側鎖ではなくデンプン主鎖を分解 |
| プロテアーゼ | ペプチド結合 | タンパク質加水分解、風味・溶解性調整 | 糖質ではなくタンパク質を標的にする |
| リパーゼ | 脂質エステル結合 | 油脂改質、風味形成、洗浄 | 糖質ではなく脂質を標的にする |
複合酵素製品の文脈では、α-ガラクトシダーゼ、リパーゼ、α-アミラーゼ、プロテアーゼ、ラクターゼが同時に挙げられることがあります。しかし、B2B工程設計では「同じカプセルに入っているか」ではなく、「どの原料画分を、どの結合で、どの順序で変換するか」を見る必要があります。β-ガラクトシダーゼの多用途性は食品・医療・工業で確立していますが、RFOやガラクトマンナン側鎖を対象にする場合はα-ガラクトシダーゼの説明に置き換えることはできません[6]。
「alpha galactosidase cofactors」という検索語は、反応に補因子が必要かを確認したい意図を反映しています。実務上は、α-ガラクトシダーゼの働きは酸化還元酵素のようにNAD、FAD、金属補酵素を中心に説明するより、基質へのアクセス、pH、温度、水分、原料マトリックス、阻害成分によって左右される加水分解反応として理解する方が適切です。食品廃棄物の酵素加水分解レビューでも、反応効率に影響する主要因としてpH、温度、基質濃度、反応時間、酵素と基質の接触性が整理されています[7]。
なお、由来生物や酵素ファミリーが異なれば、安定性、基質範囲、阻害感受性は変わります。Penicillium sp.由来α-ガラクトシダーゼでは基質特異性が研究され、Mortierella vinacea由来酵素では精製、性質、cDNAクローニングが報告されています。これは、同じα-ガラクトシダーゼという名称でも、由来により基質適合性が異なり得ることを示します[13]。
α-ガラクトシダーゼは、微生物、植物、動物、ヒトなど多様な生物に存在します。ヒトα-ガラクトシダーゼAはFabry病研究の中心であり、ヒト線維芽細胞での合成・プロセシングや変異の影響が研究されていますが、これは医療・細胞生物学の文脈です[14]。
一方、食品・飼料・工業工程では、微生物由来酵素が実装上扱いやすいことが多く、耐熱性や耐プロテアーゼ性などの性質も用途によって評価されます。Rhizopus sp.由来のGH-36 α-ガラクトシダーゼでは、プロテアーゼ抵抗性を持つ新規酵素のクローニングと発現が報告されており、消化酵素やプロセス中のタンパク質分解環境に対する安定性が研究対象になることを示しています[15]。
高温工程では、Bacillus stearothermophilus由来の耐熱性β-マンナナーゼとα-ガラクトシダーゼの精製・性質評価が報告されています。これは、α-ガラクトシダーゼが低温食品処理だけでなく、温度負荷のある多糖処理プロセスでも研究されてきたことを示します[16]。
α-ガラクトシダーゼの実装では、遊離酵素として反応液に添加するだけでなく、固定化によって再利用性や工程安定性を高める研究も行われています。Debaryomyces hansenii由来α-ガラクトシダーゼをセルロースフィルムに固定化し、オリゴ糖加水分解へ応用した研究は、液体基質中の糖質処理において固定化担体が利用できることを示しています[17]。
固定化酵素は、食品・糖質加工・発酵前処理で、酵素の回収や連続処理を考える際に有用な概念です。ただし、Enzymes.bioが供給するAlpha-Galactosidaseはオンライン販売される酵素原料であり、固定化リアクターや工程装置を製造・設計するサービスとして説明すべきではありません。固定化研究は、α-ガラクトシダーゼが多様なプロセス形態で検討されていることを理解するための科学的背景です[17]。
ヒトα-ガラクトシダーゼAは、Fabry病におけるグロボトリアオシルセラミド代謝と関連し、酵素補充療法やタンパク質変異の研究対象になっています。ヒト肝臓α-ガラクトシダーゼAでは、糖脂質およびオリゴ糖基質に対する酵素特性が研究されており、分子医学上の重要性は高い領域です[18]。
しかし、医療用酵素と食品・飼料・工業加工用酵素は、目的、規制、品質設計、使用環境が異なります。市販治療用製剤の天然基質特異性や糖鎖修飾を比較した研究もありますが、これは医薬品の同等性や作用理解に関する文脈であり、B2B加工用α-ガラクトシダーゼの治療効果を示すものではありません[19]。
α-Gal症候群についても同様です。マダニ由来酵素がα-Gal代謝に関与する可能性を扱った研究はありますが、これは免疫・アレルギー・ベクター生物学の文脈であり、食品加工用α-ガラクトシダーゼの効能表示に転用すべきではありません[20]。
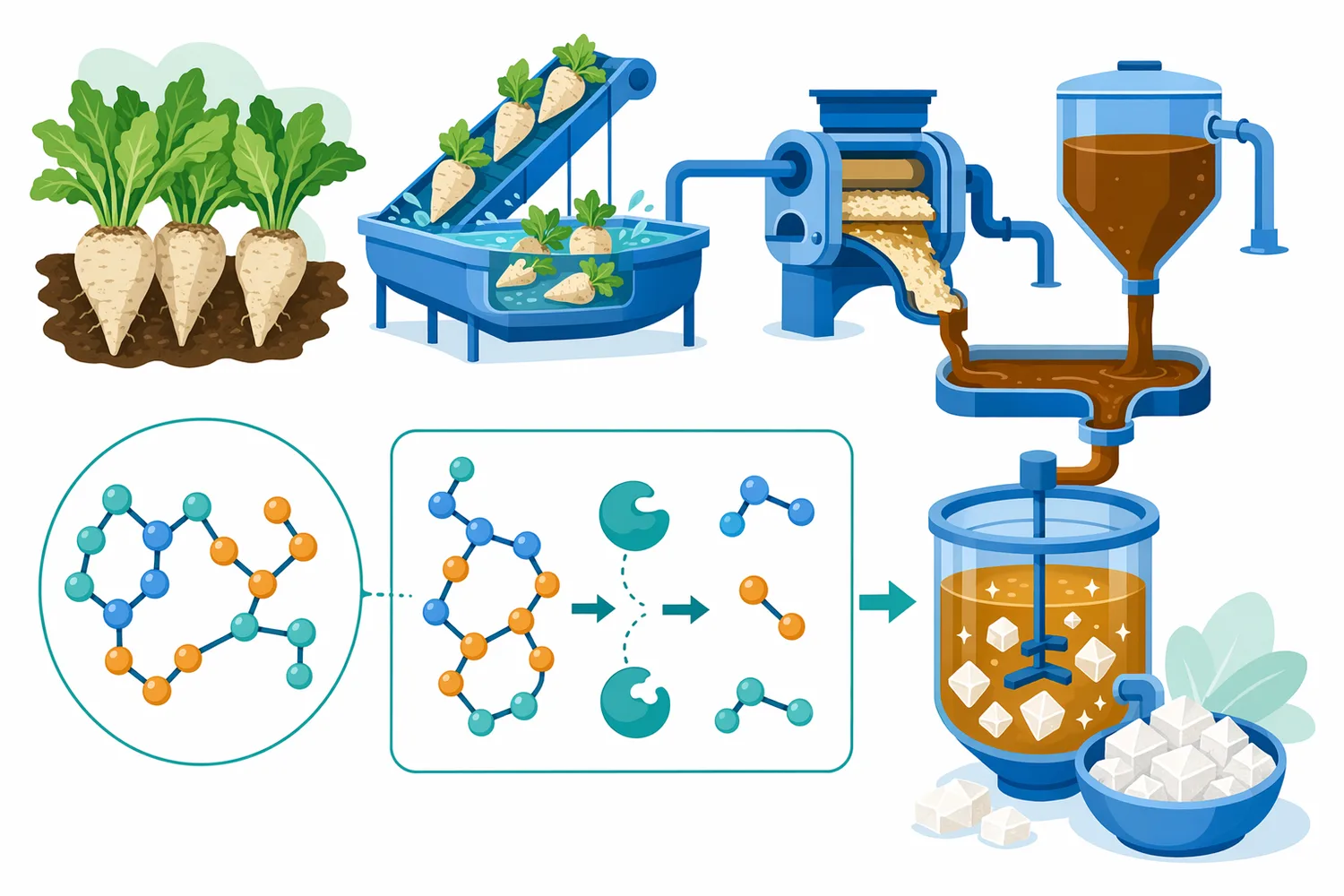
α-ガラクトシダーゼ反応では、水分を含む環境で基質と酵素が接触することが前提になります。乾燥粉末原料に酵素を混ぜただけでは、糖質が溶出せず酵素が基質にアクセスしにくい場合があります。加水、混合、保温、後段加熱、発酵接続などの順序が、同じ酵素でも結果を変えます。複合食品・副産物の酵素加水分解では、反応条件と基質構造が効率を大きく左右することが整理されています[7]。
温度は、反応速度を上げる要因である一方、過度な熱はタンパク質構造を損ないます。耐熱性α-ガラクトシダーゼの研究が行われていることは、温度負荷のある工程で酵素安定性が実用上の課題になることを示します。Thermotoga neapolitanaやBacillus stearothermophilus由来酵素の研究は、ガラクトマンナン処理や高温域の酵素利用を考える際の背景になります[9]。
pHも重要です。糖質が十分に溶出していても、pHが酵素の安定性や触媒活性に合わない場合、反応は進みにくくなります。食品原料では、豆乳、豆ペースト、糖蜜、発酵液、飼料スラリーなどでpH緩衝能や共存成分が異なるため、文献で得られた挙動をそのまますべてのマトリックスへ当てはめるのではなく、対象工程の条件に沿って理解する必要があります[7]。
Enzymes.bioは、酵素の製造業者または研究所ではなく、B2B向けのオンライン酵素供給業者です。Alpha-Galactosidaseは、植物性原料、大豆加工、食品・飼料関連工程、糖質処理を行う企業向けの酵素原料としてオンラインで直接購入できます。製品ページではAlpha-Galactosidase Enzyme CAS 9025-35-8として掲載されています。
供給単位は1kgで、オンライン注文後に処理・配送されます。CoAおよびSDSは注文時に併せて提供されるため、受領後の社内記録、安全管理、原料登録に利用できます。Enzymes.bioは工業・食品加工・B2B用途の酵素供給を行う立場であり、ヒトが直接摂取する小売用サプリメントや医療用製剤として説明するものではありません。
Alpha-Galactosidase(α-ガラクトシダーゼ)は、α-D-ガラクトシド結合を加水分解し、豆類RFO、ガラクトマンナン側鎖、糖蜜中の一部糖質などを処理する酵素です。大豆、豆類、植物性タンパク質、飼料、グアーガム、発酵基質の工程で、糖質組成、発酵性、粘度、原料利用性を調整する手段として位置づけられます[8]。
β-ガラクトシダーゼ/ラクターゼ、α-アミラーゼ、プロテアーゼ、リパーゼとは標的結合が異なるため、複合酵素の一般説明ではなく、基質構造に基づいて使い分ける必要があります。医療用ヒトα-ガラクトシダーゼやBeanoのような消費者向け経口製品の研究は参考背景にはなりますが、Enzymes.bioが供給するAlpha-GalactosidaseはB2B加工用途の酵素原料として扱うのが適切です[3]。
Enzymes.bioではAlpha-Galactosidaseを1kg単位でオンライン直接販売し、CoAとSDSは注文時に併せて提供されます。製造業者や研究機関としてではなく、B2B酵素供給業者として、植物性原料加工、食品工程、飼料工程、糖質処理に向けた酵素原料を供給します。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Alpha-Galactosidaseを購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。