Aminopeptidase Enzyme For Protein Hydrolysis は、一次加水分解で生じたペプチドのN末端側を段階的に処理し、遊離アミノ酸の増加、苦味の緩和、風味の丸みづけを支援するタンパク質加水分解向け酵素です。エンドペプチダーゼがタンパク質内部を切断してペプチドを作るのに対し、アミノペプチダーゼは生成済みペプチドを後段で整える「仕上げ酵素」として位置づけると、工程上の役割が明確になります。Enzymes.bioは本製品をB2B向け酵素としてオンラインで1 kg単位から販売しており、CoAおよびSDSは注文時に併せて提供されます。
タンパク質加水分解では、原料タンパク質を可溶化し、分子量を下げ、ペプチドやアミノ酸を含む加水分解物へ変換します。大豆、エンドウ、米、小麦、ホエイ、カゼイン、魚副産物、肉副産物、種子タンパク、微細藻類タンパクなど、原料が変わっても基本的な課題は共通しており、分解度、分子量分布、溶解性、濾過性、風味、最終用途への適合性を同時に管理する必要があります。魚廃棄物を対象にした応答曲面法による研究では、酵素加水分解条件の調整が加水分解物の収率や特性に影響することが示されており、原料ごとの工程設計が重要であることを裏づけています[1]。
一次加水分解では、中性プロテアーゼ、アルカリプロテアーゼ、アスパラギン酸プロテアーゼ、パパイン、トリプシンなどのエンドペプチダーゼが使われることが多く、これらはタンパク質鎖の内部ペプチド結合を切断して、より短いペプチドを生成します。たとえば大豆タンパク質を対象にした酵素加水分解研究では、反応条件と時間により加水分解の進行が変化し、ラボスケールと半工業スケールでの挙動検討が行われています[2]。しかし、内部切断だけでは、疎水性アミノ酸を含む短鎖ペプチドや、後味の粗さにつながるペプチドが残る場合があります。
アミノペプチダーゼは、一次加水分解後に増えたペプチドを基質として利用し、N末端側からアミノ酸を順に遊離します。これにより、単に分子量を下げるだけでなく、ペプチド末端構造、遊離アミノ酸比率、苦味の出方、うま味・甘味・後味のバランスを調整しやすくなります。Thermomyces lanuginosus由来の耐熱性ロイシンアミノペプチダーゼを大豆タンパク質加水分解に応用した研究は、アミノペプチダーゼが植物タンパク加水分解物の後処理酵素として検討されていることを示す代表的な例です[3]。
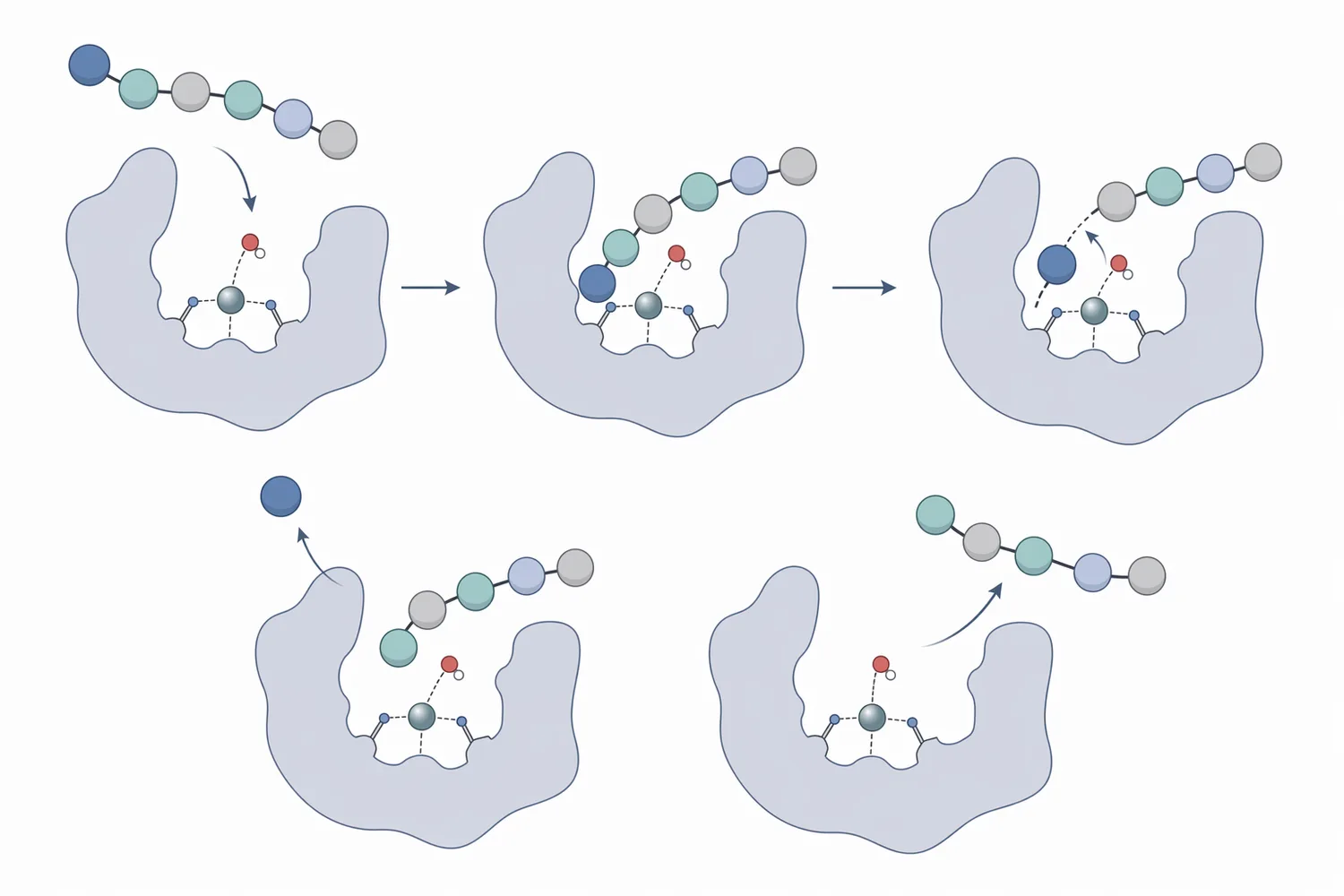
タンパク質は、アミノ酸がペプチド結合で連結した高分子です。タンパク質加水分解で重要なのは、どの位置のペプチド結合を切るかです。エンドペプチダーゼはタンパク質鎖の内部を切断するため、長いタンパク質を多数のペプチドへ短時間で分割しやすい一方、生成物は配列や長さが多様な混合物になります。これに対して、アミノペプチダーゼはエキソペプチダーゼの一種であり、ペプチドのN末端からアミノ酸を一つずつ外していく点が特徴です。PepNなどのアミノペプチダーゼと他のエキソペプチダーゼを組み合わせた研究では、末端からの段階的処理がペプチド組成調整に利用できることが示されています[4]。
| 項目 | エンドペプチダーゼ | アミノペプチダーゼ |
|---|---|---|
| 主な切断位置 | タンパク質鎖・ペプチド鎖の内部 | ペプチドのN末端側 |
| 工程上の役割 | 一次加水分解、可溶化、分子量低下 | 二次加水分解、末端処理、風味調整 |
| 主な生成物 | 多様な長さのペプチド | 遊離アミノ酸、末端が変化したペプチド |
| 実務上の狙い | 原料タンパク質を分解し、後工程に進めやすくする | 苦味低減、遊離アミノ酸増加、味質の丸みづけ |
| 単独使用の位置づけ | 主分解酵素として使いやすい | 仕上げ酵素として使いやすい |
この違いは、実務上きわめて重要です。アミノペプチダーゼを最初から未分解タンパク質に使っても、基質となるN末端の数が限られるため、エンドペプチダーゼほど大きく分子量を下げる働きは期待しにくくなります。先に内部切断でペプチド数を増やし、その後にアミノペプチダーゼで末端処理を行うほうが、酵素の役割分担として合理的です。Aspergillus niger由来アスパラギン酸プロテアーゼを用いた大豆タンパク質分解の研究でも、プロテアーゼによる一次的なタンパク質分解が加水分解物特性を左右することが示されています[5]。
アミノペプチダーゼには複数の酵素ファミリーがあり、由来生物や構造によって触媒機構は異なります。多くのアミノペプチダーゼでは、活性部位の金属イオン、配位残基、酸塩基触媒に関わるアミノ酸残基が、ペプチド結合の加水分解に関与します。Aeromonas proteolytica由来アミノペプチダーゼの速度論、分光、X線結晶構造解析を組み合わせた研究では、活性部位構造と触媒機能の関係が詳細に検討されています[6]。
ただし、基礎研究で明らかになった特定酵素の構造や金属依存性を、商用製剤すべてにそのまま当てはめることはできません。食品加工・発酵原料・栄養素材向けの産業用途では、精製酵素単体の構造情報よりも、実際の原料タンパク質、一次加水分解の状態、pH、温度、反応時間、後処理との組み合わせが性能を大きく左右します。したがって、アミノペプチダーゼは「特定の健康機能を与える成分」ではなく、「ペプチド末端を変化させる工程用酵素」として理解するのが適切です。
タンパク質加水分解物の苦味は、単純に「分解が足りない」または「分解しすぎた」という一要因だけで説明できません。疎水性アミノ酸を含むペプチド、ペプチドの長さ、配列、N末端・C末端の状態、遊離アミノ酸との比率、原料由来の香気成分が組み合わさって官能品質が決まります。種子タンパクや副産物タンパクから酵素加水分解物を作る研究では、加水分解条件により抗酸化性などの機能指標だけでなく、生成ペプチドの性質も変化することが報告されています[7]。
アミノペプチダーゼの利点は、苦味ペプチドを一気に別物へ変えるのではなく、末端から少しずつアミノ酸を外す点にあります。N末端が変わることで、苦味受容に関わるペプチド構造が変化し、同時に遊離アミノ酸が増加します。遊離グルタミン酸やアスパラギン酸はうま味・酸味に、グリシンやアラニンは甘味に、ロイシンやフェニルアラニンなどは苦味に関与し得るため、単にアミノ酸量を増やせば必ず好ましい風味になるわけではありません。二酵素加水分解を用いたクルミ搾りかすタンパク質の研究では、酵素の組み合わせが加水分解物の組成や性質に影響することが示されています[8]。
そのため、アミノペプチダーゼは「苦味をゼロにする酵素」ではなく、「一次加水分解物の末端構造を変え、風味を調整する酵素」と表現するのが正確です。苦味低減を狙う場合でも、一次加水分解でどの程度ペプチドが生成しているか、疎水性ペプチドがどの程度残っているか、最終用途が調味、栄養、発酵窒素源、ペットフード、培地成分のどれかによって、望ましい処理の深さは変わります。超音波併用を含むカボチャ種子タンパク質加水分解の研究でも、処理条件によって物理化学的性質と抗酸化活性が変わることが示されており、加水分解は一律の反応ではなく設計対象であることがわかります[9]。
植物タンパク加水分解物では、豆臭、青臭さ、渋味、苦味、粉っぽさが課題になることがあります。大豆タンパク、エンドウタンパク、小麦グルテン、米タンパク、ヒマワリ、トウモロコシ、ソルガムなどでは、原料自体の難溶性や疎水性領域、前処理による変性状態が加水分解挙動に影響します。ヒマワリ副産物や種子タンパクを対象にした研究では、酵素加水分解により生理活性ペプチドや抗酸化性を持つ画分の生成が検討されています[7]。
大豆タンパク質では、一次プロテアーゼで可溶化と低分子化を進めた後、アミノペプチダーゼで末端処理を行う流れが考えられます。Thermomyces lanuginosus由来ロイシンアミノペプチダーゼの大豆タンパク質加水分解への応用研究は、植物タンパク加水分解物の品質調整にアミノペプチダーゼが関与し得ることを示しています[3]。調味ベース、植物性プロテイン飲料、発酵原料、培地用窒素源では、溶解性と風味の両方が求められるため、アミノペプチダーゼの後段使用が工程上の選択肢になります。
ホエイタンパクやカゼインは、栄養素材、スポーツ栄養、医療・介護食向け素材、乳由来ペプチド素材に広く使われます。乳タンパクは高栄養価である一方、加水分解を深く進めると苦味が出やすく、特に高分解度品では官能品質が製品設計の制約になります。小麦、エンドウ、米、ホエイの消化性を比較したin vitro研究では、タンパク質源によって消化挙動が異なることが示されており、乳タンパクと植物タンパクを同じ条件で扱えないことがわかります[10]。
アミノペプチダーゼは、乳タンパク加水分解物中の苦味ペプチドを後段で処理する目的に適しています。特にカゼイン由来ペプチドは疎水性配列を含むことがあり、エンドペプチダーゼだけでは苦味が残る場合があります。PepNのようなアミノペプチダーゼと他のエキソペプチダーゼを組み合わせた研究は、ペプチド末端処理が乳タンパク系加水分解物の風味調整に応用されてきた背景を示しています[4]。
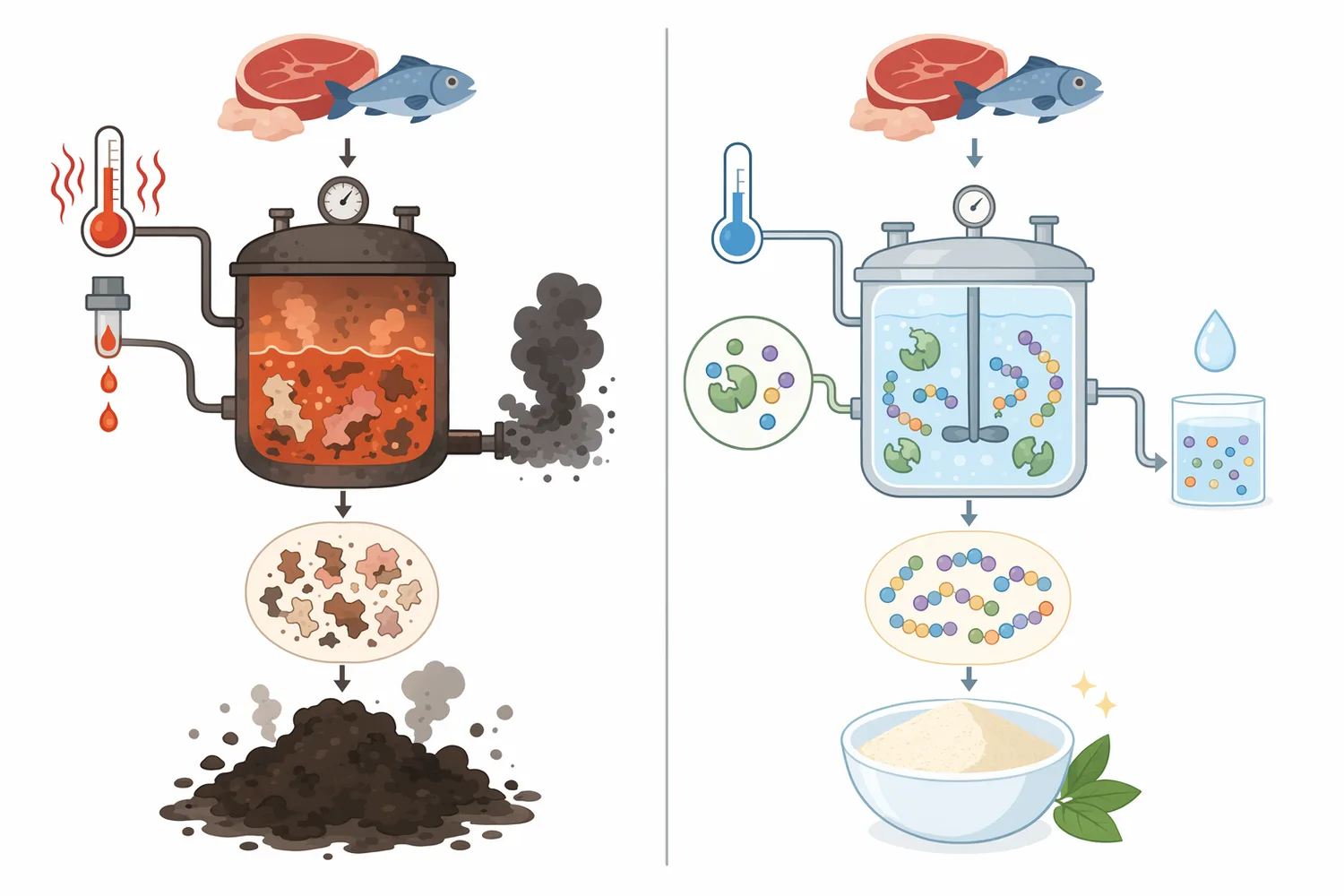
魚の皮、骨、頭部、内臓、端材、食肉加工副産物は、タンパク質を多く含む一方、そのままでは用途が限られます。酵素加水分解により、これらをペプチド、アミノ酸、可溶性窒素を含む素材へ変換すれば、調味、飼料、ペットフード、発酵原料、バイオプロセス用窒素源などへの利用可能性が広がります。魚副産物由来酵素と加水分解物に関するレビューでは、水産副産物を循環型の生理活性加水分解物へ変換する考え方が整理されています[11]。
魚骨タンパク質のトリプシン加水分解物からACE阻害ペプチドを得た研究のように、魚由来タンパクは特定ペプチド探索の出発原料にもなります。ただし、このような研究は特定原料、特定酵素、特定ペプチド配列に基づくものであり、アミノペプチダーゼ製剤そのものの健康機能を意味するものではありません[12]。アミノペプチダーゼの実務的価値は、こうした加水分解物をさらに加工し、遊離アミノ酸とペプチド末端構造を調整する点にあります。
微細藻類タンパクは、持続可能なタンパク源として注目されており、細胞破砕、抽出、酵素加水分解、分離精製を組み合わせてペプチド素材へ加工されます。微細藻類タンパク抽出物の酵素加水分解を最適化した研究では、加水分解が機能性・構造特性の改善に関連することが検討されています[13]。こうした新規タンパクでは、原料細胞壁、色素、脂質、灰分、原料臭が工程に影響するため、単にプロテアーゼを加えるだけでは品質を制御しにくい場合があります。
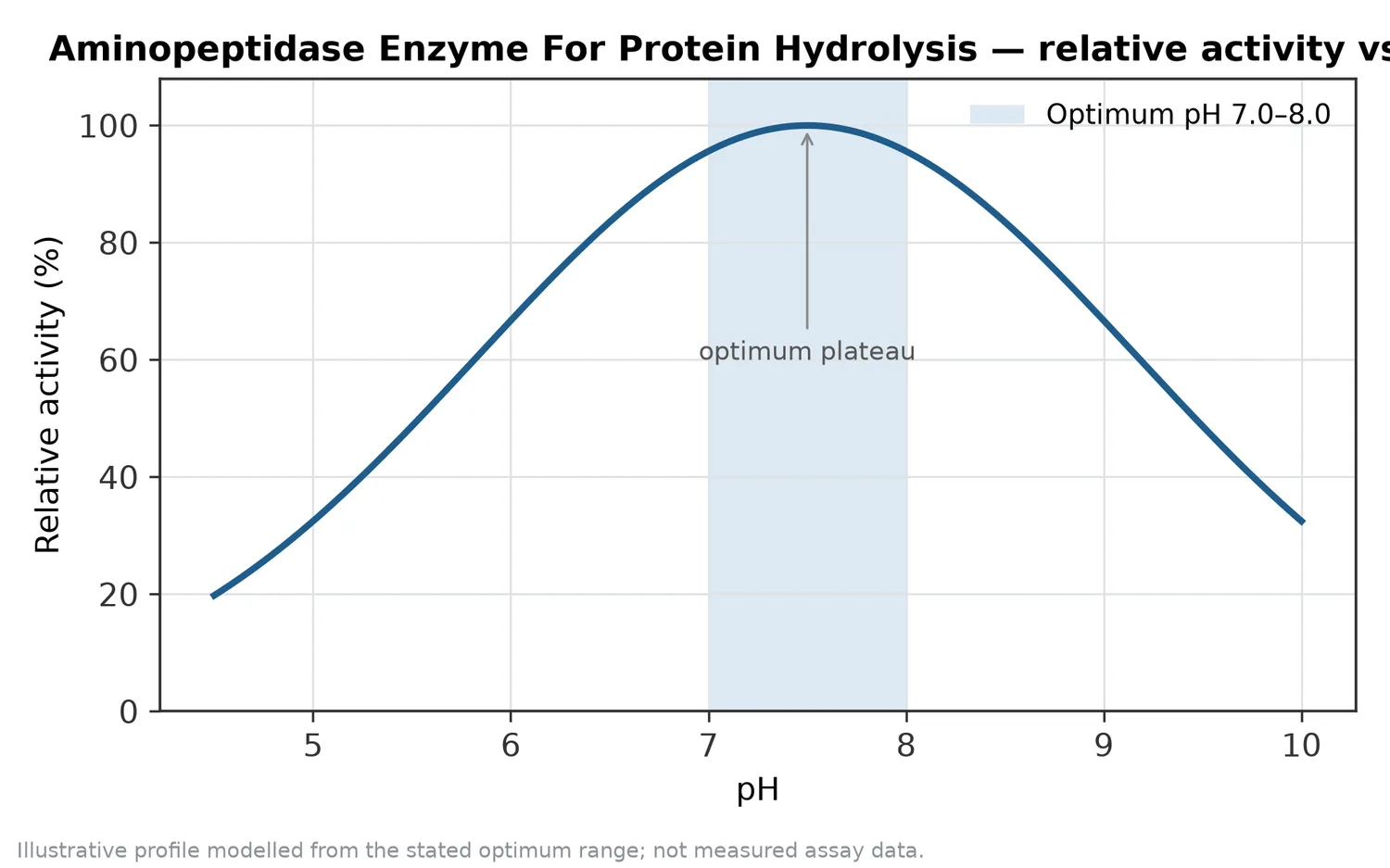
アミノペプチダーゼは、新規タンパクの一次加水分解物に対しても、末端処理と遊離アミノ酸増加を通じて風味や溶解性の調整に使える可能性があります。ただし、新規タンパクでは原料ロット、前処理、抽出方法の影響が大きいため、アミノペプチダーゼの効果は原料タンパクの構造と一次加水分解の状態に強く依存します。これは微細藻類に限らず、植物副産物、発酵菌体、昆虫タンパク、単細胞タンパクにも共通する工程上の考え方です。
アミノペプチダーゼを工程に入れる典型的な流れは、原料分散、pH調整、一次加水分解、必要に応じた固液分離、二次加水分解、酵素失活、濾過・濃縮・乾燥です。ここでアミノペプチダーゼは、一次加水分解で十分なペプチド基質が形成された後に導入されます。ペリラ粕タンパク質の酵素加水分解を最適化した研究では、高い加水分解度と抗酸化活性を持つ加水分解物を得るために反応条件の最適化が検討されており、加水分解工程では条件設定が最終特性に直結することが示されています[14]。
二段階処理の利点は、役割分担が明確になることです。一次酵素はタンパク質を崩して可溶化し、二次酵素であるアミノペプチダーゼは生成済みペプチドを末端から整えます。絹セリシン加水分解物の応答曲面最適化研究のように、反応条件と目的指標の関係を整理するアプローチは、タンパク質加水分解物の品質設計に広く用いられています[15]。アミノペプチダーゼを使う場合も、目的が苦味低減なのか、遊離アミノ酸増加なのか、発酵適性改善なのかによって、処理の深さを変える必要があります。
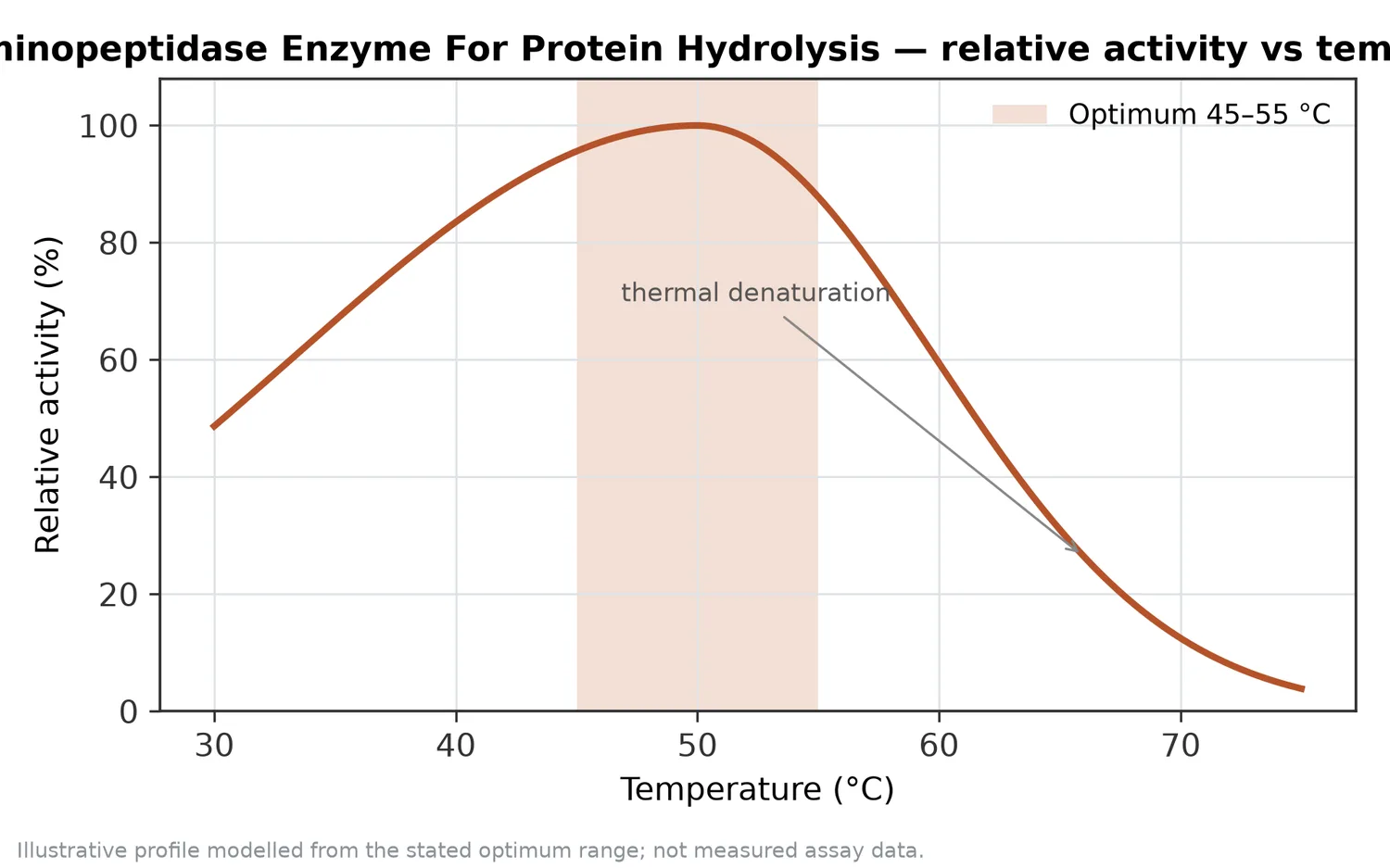
過度な処理は常に望ましいわけではありません。遊離アミノ酸が増えすぎると、うま味が強くなる一方で、苦味、酸味、金属味、後味の残り方が変化する場合があります。また、ペプチドを短くしすぎると、目的とするペプチド画分の分子量分布や機能性指標が変わる可能性があります。サトウキビストローなどのリグノセルロース分解研究でも示されるように、酵素反応では基質構造、酵素の組み合わせ、反応時間の相互作用が成果を左右しますが、タンパク質加水分解でも同様に「長く反応させればよい」という単純な設計は成り立ちません[16]。
| 用途領域 | 一次加水分解での主課題 | アミノペプチダーゼ導入の狙い | 留意すべき点 |
|---|---|---|---|
| 調味素材・うま味ベース | 苦味、渋味、後味の粗さ | 遊離アミノ酸増加、味の丸みづけ | 過度な遊離アミノ酸化で味が重くなる場合がある |
| 栄養素材・プロテイン加水分解物 | 高分解度化に伴う苦味 | 苦味ペプチドの末端処理 | 健康機能を酵素自体に帰属させない |
| 発酵用窒素源 | 微生物が利用しやすい窒素形態の不足 | 短鎖ペプチド・遊離アミノ酸の増加 | 微生物種ごとに好む窒素形態が異なる |
| 魚・肉副産物の価値化 | 原料臭、難利用部位、低い可溶性 | 可溶性窒素と風味の調整 | 脂質酸化臭やミネラル由来味は別管理が必要 |
| 植物タンパク素材 | 豆臭、青臭さ、苦味、沈殿 | ペプチド末端処理と溶解性改善の補助 | 原料前処理の影響が大きい |
発酵用窒素源として考える場合、アミノペプチダーゼの価値は、微生物が利用しやすい窒素形態を増やす点にあります。タンパク質リッチな産業廃棄物を同時加水分解する研究では、タンパク質性副産物を酵素処理して有用な加水分解物へ変換する可能性が示されています[17]。この用途では、官能品質だけでなく、可溶性窒素、ペプチド長、発酵時の泡立ち、濾過性、滅菌後の沈殿なども実務上の評価対象になります。
調味素材では、苦味低減とうま味付与のバランスが重要です。アミノペプチダーゼにより遊離アミノ酸が増えると、味の立ち上がりや後味が変化しますが、目的の風味プロファイルは用途ごとに異なります。たとえば、スープベースではうま味とコクが重視され、プロテイン飲料では苦味と後味の軽さが重視され、ペットフードでは嗜好性と加工安定性が重視されます。赤海藻寒天抽出後の産業残渣を酵素加水分解した研究では、産業残渣も条件設計により抽出物特性が変わることが示されており、副産物利用では原料由来の特性を踏まえた処理が不可欠です[18]。
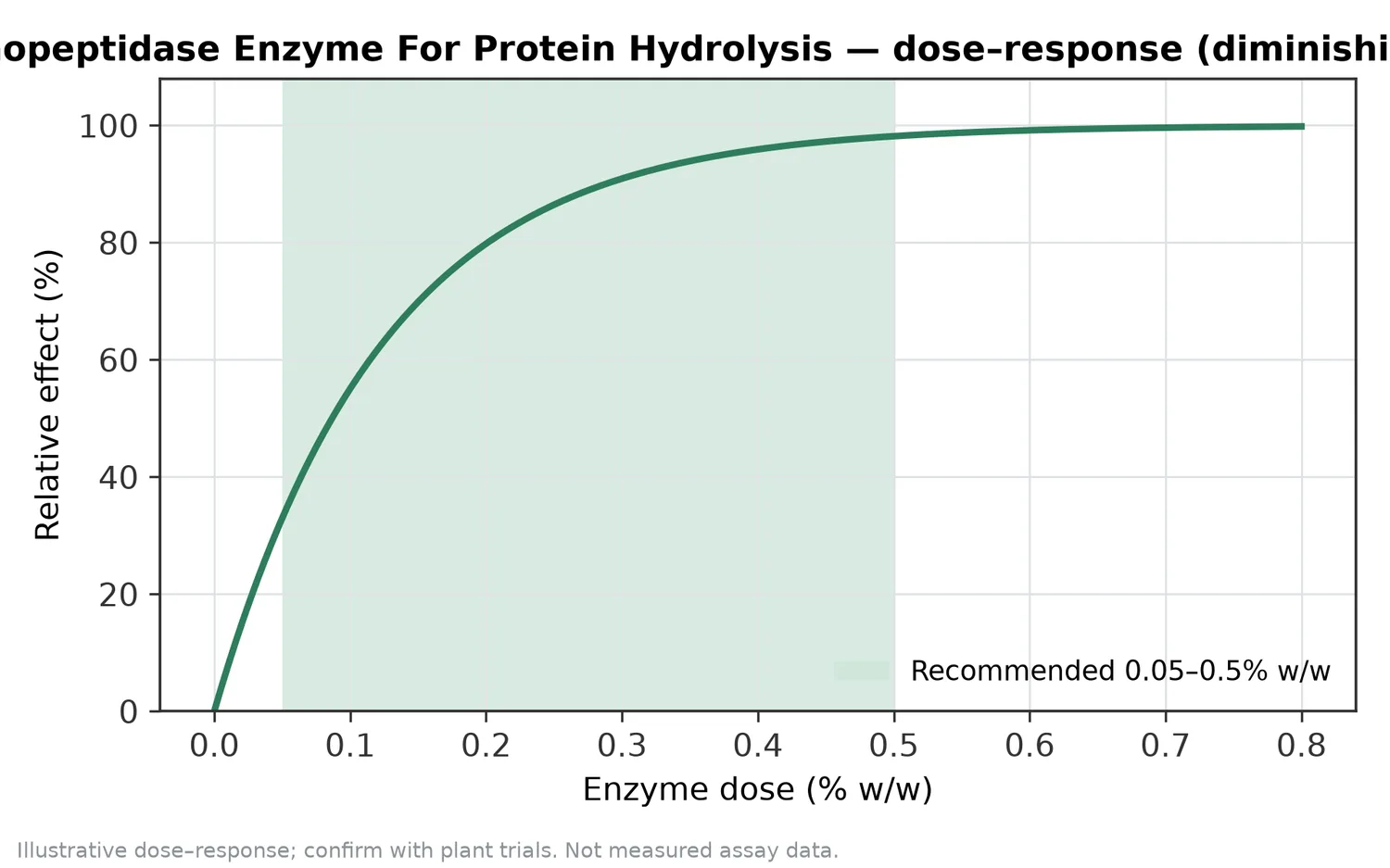
タンパク質加水分解物に関する研究では、抗酸化活性、ACE阻害活性、ROS捕捉活性などの指標がしばしば測定されます。たとえば、サーモン骨タンパク質のトリプシン加水分解物からACE阻害ペプチドを得た研究や、絹セリシン加水分解物のROS捕捉活性を最適化した研究は、加水分解物が機能性素材開発の出発点になり得ることを示しています[12][15]。
しかし、これらの機能性は、原料タンパク質、酵素選択、加水分解条件、ペプチド配列、分離画分、評価系に依存します。アミノペプチダーゼはペプチド末端を処理する酵素であり、最終製品の健康効果を直接保証するものではありません。アミノペプチダーゼ処理により特定の生理活性ペプチドが増える場合もあれば、逆に目的ペプチドがさらに分解される場合もあります。したがって、Enzymes.bioの本製品を説明する際は、「機能性を付与する有効成分」ではなく、「タンパク質加水分解工程でペプチド組成と風味を調整する加工用酵素」と表現するのが科学的に妥当です。
Enzymes.bioは酵素の供給業者であり、Aminopeptidase Enzyme For Protein Hydrolysisをタンパク質加水分解向け酵素としてオンライン販売しています。本製品は1 kg単位で購入でき、B2Bの食品加工・産業加工用途を想定した酵素です。注文時にはCoAおよびSDSが併せて提供されるため、受領後の社内受入、保管、工程投入に必要な基本文書を確認できます。
Enzymes.bioのタンパク質加水分解向け酵素カテゴリには、アミノペプチダーゼ以外にも、タンパク質を一次分解する各種プロテアーゼが含まれます。アミノペプチダーゼは、それらの一次分解酵素と競合するものではなく、むしろ後段で補完的に使う酵素です。一次分解でペプチド基質を作り、二次分解で遊離アミノ酸と末端構造を調整するという考え方により、タンパク質加水分解物の風味、溶解性、発酵適性、後工程適性を設計しやすくなります。
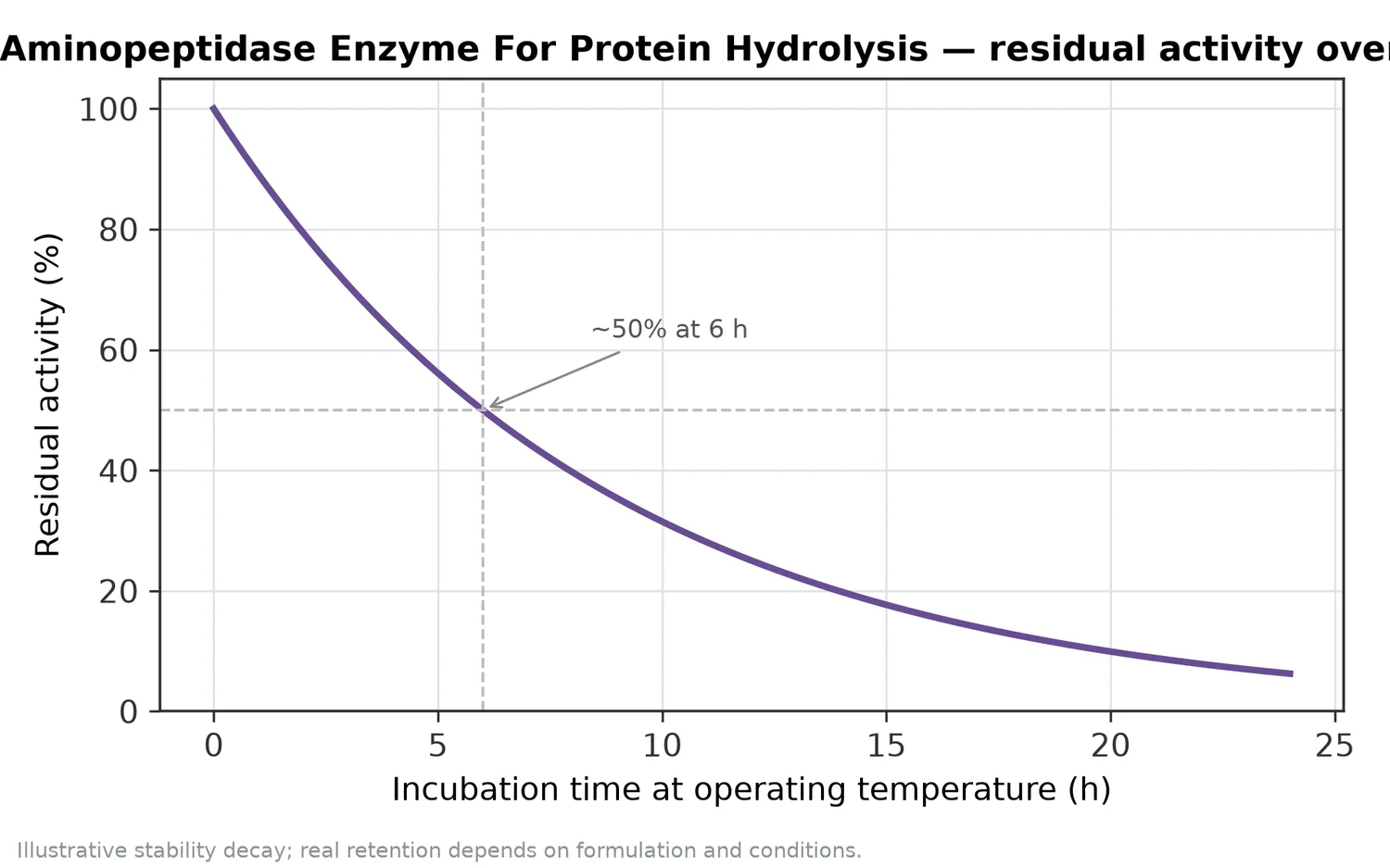
Aminopeptidase Enzyme For Protein Hydrolysisは、タンパク質加水分解の主分解酵素というより、一次加水分解後のペプチド混合物を仕上げる酵素です。N末端側からアミノ酸を段階的に遊離することで、遊離アミノ酸量を増やし、苦味ペプチドの末端構造を変え、風味の丸みづけや後工程適性の改善に寄与します。特に、植物タンパク加水分解物、乳タンパク加水分解物、魚・肉副産物由来ペプチド、発酵用窒素源、調味素材の設計で有用です。
一方で、アミノペプチダーゼは最終製品の健康効果を保証するものではなく、特定の生理活性ペプチド生成を常に促進するものでもありません。研究文献は、酵素加水分解がタンパク質副産物の価値化、ペプチド素材開発、抗酸化性やACE阻害活性などの探索に有効であることを示していますが、その成果は原料、酵素、条件、分離画分に依存します[11][12]。本製品の価値は、タンパク質加水分解物の品質を工程内で調整し、苦味、遊離アミノ酸、ペプチド組成、用途適性を制御しやすくする点にあります。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Aminopeptidase Enzyme For Protein Hydrolysisを購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。