Aminopeptidase Enzyme For Protein Hydrolysis 是用於蛋白質水解流程的胺肽酶,主要應用在水解後段,從胜肽的 N 端逐步釋放胺基酸,協助調整水解物的風味、苦味與游離胺基酸輪廓。
它通常不是取代初段蛋白酶,而是與內切型蛋白酶搭配,用於植物蛋白水解物、調味基底、營養配方、膠原與動物蛋白水解物等應用。
Enzymes.bio 作為酵素供應商,提供 1kg 單位線上購買;CoA 與 SDS 會隨訂單一併提供。
Aminopeptidase Enzyme For Protein Hydrolysis 指的是可用於蛋白水解流程的胺肽酶。從製程定位來看,胺肽酶屬於外切型蛋白水解酵素,主要從胜肽鏈的胺基端,也就是 N 端,逐步移除單一胺基酸或促進末端胜肽縮短。這與常見的內切型蛋白酶不同;內切型蛋白酶多在蛋白質鏈內部切割,快速降低分子量並產生多種長短不一的胜肽,而胺肽酶更常被用在後段,調整已形成的胜肽組成與游離胺基酸比例。食品生物技術文獻也將微生物酵素視為食品加工中具有特異性的生物催化工具,可用於蛋白質、澱粉、脂質等原料的轉化與品質調整[1]。
在蛋白水解物開發中,胺肽酶的價值通常不是「把完整蛋白質最快切開」,而是協助把前段水解產生的小胜肽進一步修飾。當水解液中累積較多短胜肽時,胺肽酶可持續作用於其 N 端,改變末端胺基酸暴露情形、降低部分苦味胜肽的感官影響,並提升游離胺基酸的可用比例。這類從胜肽前驅物逐步修剪的概念,也可在 ERAP1 等胺肽酶研究中看到:其結構研究顯示,胺肽酶透過活性位點與底物結合口袋辨識胜肽,並進行逐步修剪,而非隨機破壞整條蛋白鏈[2]。
Enzymes.bio 不是製造商,也不是檢測實驗室;本文定位為 B2B 技術教育文件,協助食品加工、蛋白水解物、調味基底與營養配方開發者理解胺肽酶的應用邏輯與科學基礎。Enzymes.bio 的蛋白水解酵素分類中,列有供蛋白水解應用使用的酵素產品,Aminopeptidase Enzyme For Protein Hydrolysis 可作為此類流程中的後段修飾酵素之一。
蛋白質被水解後,會形成不同長度、不同胺基酸組成與不同疏水性的胜肽。當水解程度提高時,某些疏水性胜肽可能累積並造成明顯苦味,這是植物蛋白水解物、乳蛋白水解物、膠原水解物、肉類副產物水解物與調味基底常見的開發障礙。胺肽酶可從 N 端逐步去除胺基酸,使部分苦味胜肽進一步縮短或改變末端組成,因此常被視為高水解度產品的後段去苦工具之一;其效果並非遮蔽味道,而是從水解物組成層面改變風味來源。
此機制的關鍵在於「末端修飾」。許多苦味胜肽的感官表現與疏水性胺基酸暴露、胜肽長度與序列有關;若胺肽酶能移除 N 端特定胺基酸,可能降低胜肽與味覺受體的互動強度,或將苦味胜肽轉化為較短、較不苦的片段。研究顯示,不同胺肽酶的 S1 specificity pocket,也就是辨識 N 端第一個胺基酸側鏈的口袋,會決定其偏好的底物類型,這解釋了為何不同胺肽酶在去苦與風味修飾上的表現可能不同[3]。
在鹹味調味基底、湯底、醬汁、植物肉風味系統與營養配方中,游離胺基酸會影響鮮味、甜味、苦味、回甘、厚味與背景風味。胺肽酶的作用方式正好符合這類需求:它不只是把蛋白質變小,而是把胜肽端點的胺基酸逐步釋放出來,讓水解液中的游離胺基酸比例上升。對於希望建立酵母抽出物替代方案、植物蛋白調味基底或肉味反應前驅物的開發者而言,這種後段釋放胺基酸的能力具有實務價值。
不過,胺肽酶不應被描述為能「直接創造」特定肉味、鮮味或發酵味。風味最終仍取決於蛋白來源、前段蛋白酶切割模式、反應條件、熱處理、還原糖、鹽度、酸度與配方背景。胺肽酶提供的是可調整的胜肽與胺基酸組成,而不是固定風味配方。以白胺酸胺肽酶為例,結構研究顯示其活性位點可容納不同底物並展現廣泛底物適應性,這支持其在蛋白水解物後段調整中具有彈性,但也表示實際結果會受到底物組成影響[4]。
工業蛋白水解常見策略是二段式或多段式設計:第一段使用內切型蛋白酶快速打開蛋白質結構、降低分子量並提高可溶性;第二段再使用外切型酵素,例如胺肽酶,針對短胜肽做風味與游離胺基酸精修。這樣的分工能讓水解流程更可控,也能避免單一酵素同時承擔「快速降解」與「精細風味修飾」兩種目標。
這種分段邏輯也符合酵素工業應用的基本原則:不同酵素的結構、底物辨識能力與反應位置不同,適合放在不同製程階段。細菌 aminopeptidase P 的研究即指出,特定胺肽酶可針對含脯胺酸相關序列進行處理,並具有與食品及生物技術相關的工業意義;這說明胺肽酶不是單一功能的通用工具,而是一組可依底物需求選擇與配置的外切型酵素類群[5]。
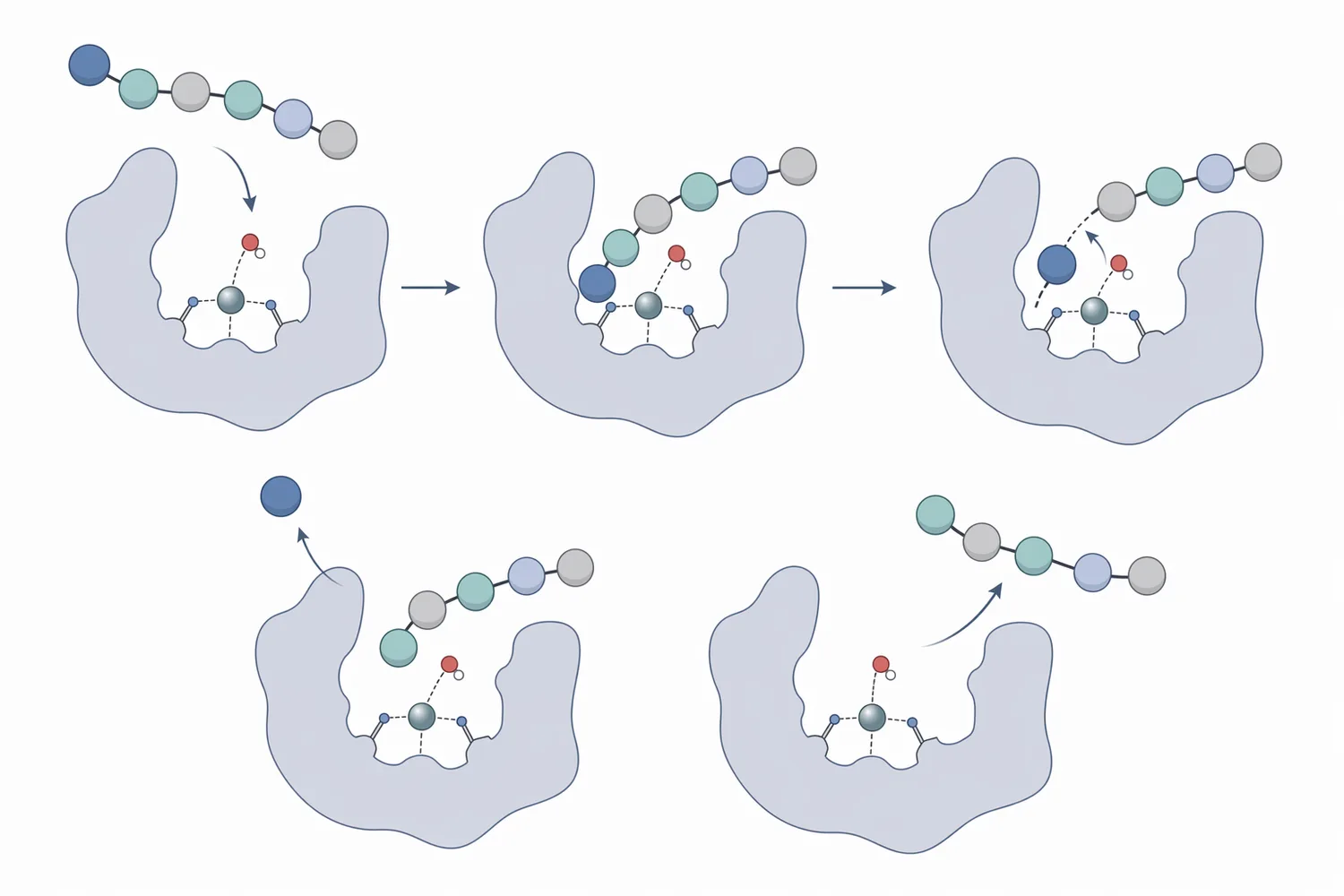
蛋白質可視為由多個胺基酸串接而成的長鏈。內切型蛋白酶像是在鏈中間剪開,快速產生多個胜肽片段;胺肽酶則像是從鏈的一端逐格拆解,主要作用於 N 端。當 N 端胺基酸被辨識並定位到活性位點後,酵素會促進胜肽鍵水解,使該胺基酸釋放為游離形式,剩餘胜肽則露出新的 N 端,可能再次進入下一輪反應。
這種逐步修剪機制使胺肽酶特別適合處理「已經被切短」的胜肽,而不是完整、折疊緊密的大分子蛋白。ERAP1 的結構研究顯示,胺肽酶可透過構形變化形成適合胜肽修剪的反應空間,並將底物導向活性位點;雖然 ERAP1 屬於免疫抗原胜肽加工研究脈絡,但其結構概念有助於理解胺肽酶為何需要可進入、可定位的胜肽底物[2]。
胺肽酶的應用表現很大程度取決於底物特異性。不同胺肽酶對 N 端胺基酸的偏好不同,有些偏好疏水性胺基酸,有些能處理酸性或鹼性胺基酸,有些則對脯胺酸鄰近序列更具意義。這些差異並非單靠反應條件能完全改變,而是與酵素活性位點附近的結構口袋、電荷分布與空間限制有關。
S1 口袋研究清楚顯示,胺肽酶辨識 N 端胺基酸側鏈的方式會影響其生成抗原胜肽或修剪胜肽的能力;換到食品與蛋白水解場景,這代表不同胺肽酶會對水解物中不同胜肽族群產生不同影響[3]。因此,Aminopeptidase Enzyme For Protein Hydrolysis 的合理定位,是協助調整水解物組成與風味輪廓,而不是保證所有蛋白原料都產生相同結果。
近年 M61 family aminopeptidase 的晶體結構研究也顯示,某些胺肽酶可具備廣泛底物特異性,並在酸性胺基酸回收中扮演關鍵角色;這進一步說明胺肽酶類群的功能差異相當大,從食品蛋白水解角度看,底物範圍與末端胺基酸偏好會直接影響水解物的最終組成[6]。
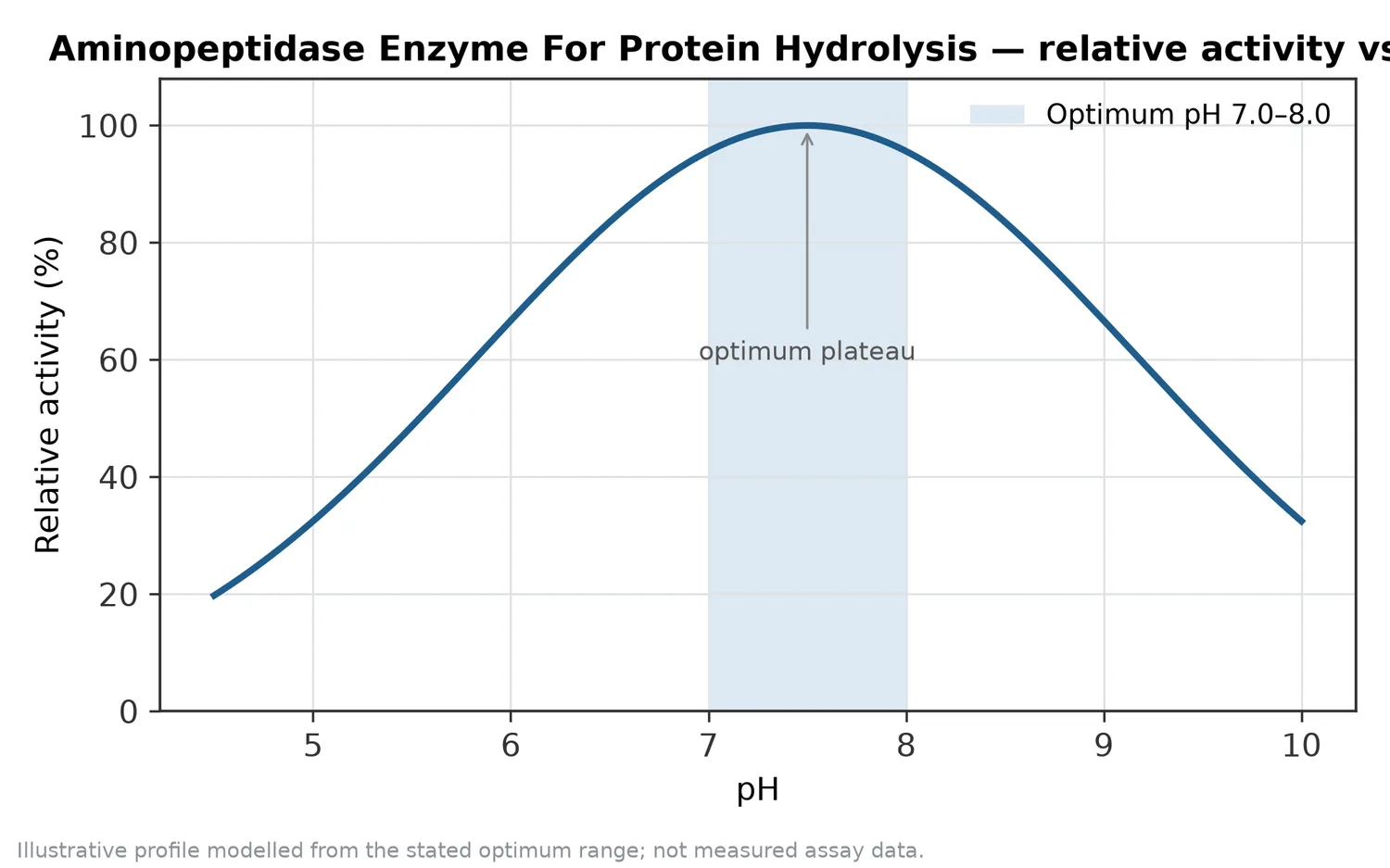
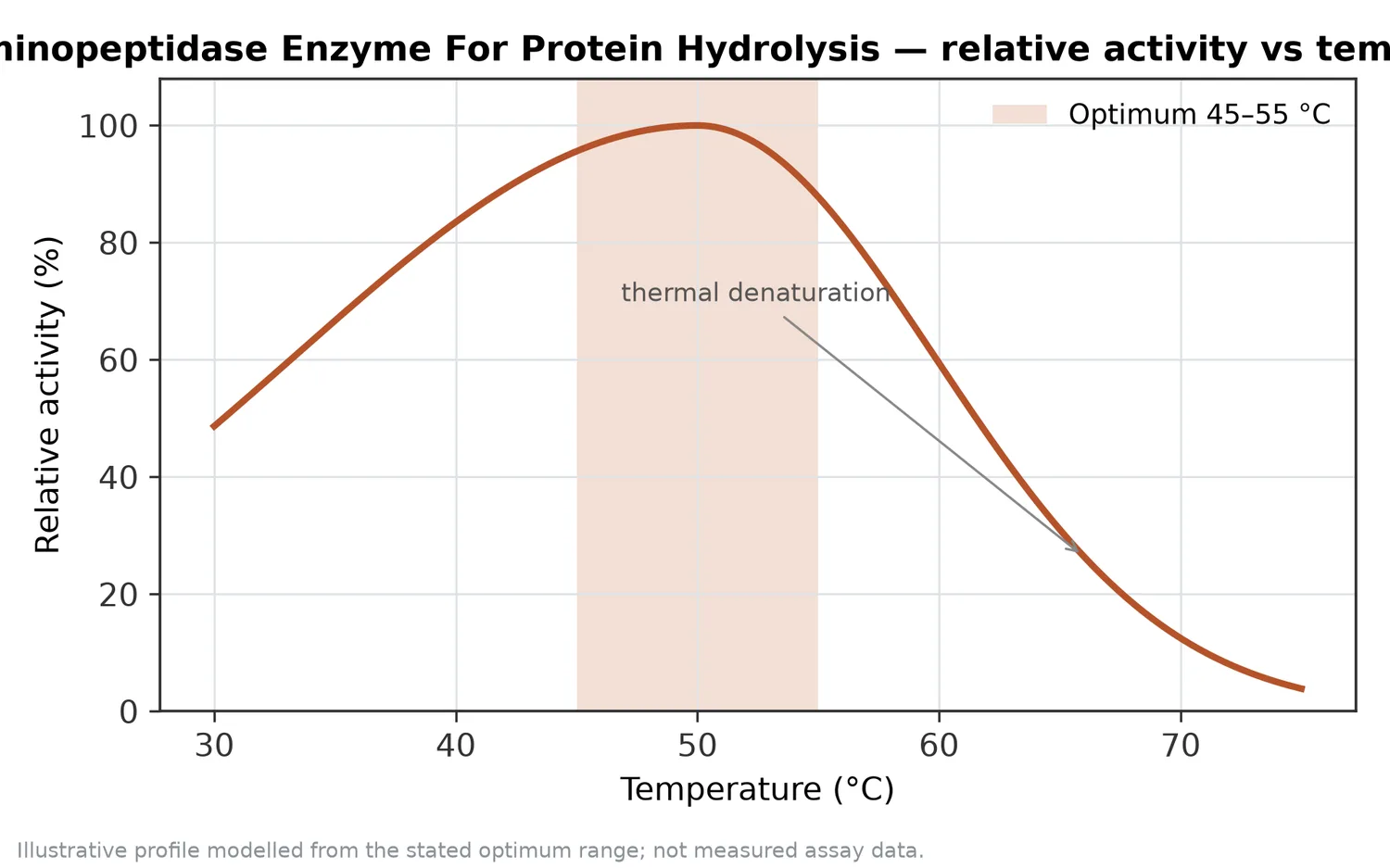
許多胺肽酶屬於金屬蛋白酶,活性位點中金屬離子會協助定位水分子、穩定反應中間態,並促進胜肽鍵水解。這不代表使用者需要在應用文件中聚焦於分析方法或活性單位定義;更重要的是理解,胺肽酶的反應對環境條件敏感,pH、溫度、鹽度、基質濃度與配方成分都可能影響酵素構形、底物可及性與反應效率。
白胺酸胺肽酶研究指出,酵素的活性位點結構與底物結合方式可解釋其廣泛底物特異性與立體選擇性;換言之,即使同樣稱為 aminopeptidase,不同來源或不同家族的酵素也可能呈現不同的底物偏好與加工結果[4]。對實務端而言,這意味著製程設計應把胺肽酶視為「需與原料及前段水解條件共同匹配」的生物催化工具。
蛋白水解流程通常不是單一酵素即可達成所有目標。內切型蛋白酶、胺肽酶與其他外切型酵素在切割位置、反應階段與產品效果上各有角色。下表以製程功能角度整理差異,協助理解 Aminopeptidase Enzyme For Protein Hydrolysis 最適合放在哪個位置。
| 酵素類型 | 主要切割位置 | 常見製程角色 | 對水解物的主要影響 | 使用上的重點 |
|---|---|---|---|---|
| 內切型蛋白酶 | 蛋白質或長胜肽內部 | 初段水解、快速降低分子量 | 增加可溶性胜肽、降低黏度、建立水解基礎 | 適合先打開蛋白結構,但可能產生苦味胜肽 |
| 胺肽酶 | 胜肽 N 端 | 後段精修、二段水解、去苦與風味調整 | 提高游離胺基酸、縮短部分胜肽、改善風味輪廓 | 需要前段水解提供可作用短胜肽 |
| 羧肽酶類外切酵素 | 胜肽 C 端 | 末端修飾、特定胜肽調整 | 改變 C 端胺基酸釋放與胜肽組成 | 與胺肽酶方向不同,可依目標搭配 |
| 複合蛋白酶系統 | 多個切割位置 | 多段或複合水解 | 同時影響水解度、分子量分布與風味 | 彈性高,但需要控制反應終點 |
這張表的核心訊息是:胺肽酶並不是初段水解的唯一選擇,而是後段精修工具。食品酵素應用文獻指出,微生物酵素在食品生物技術中常因其專一性與加工條件相容性被選用;然而,酵素功能必須與原料特性及製程目標配合,才能形成穩定的加工結果[1]。
植物蛋白水解物常見原料包括大豆、豌豆、小麥、米蛋白與其他豆類蛋白。這些原料在高蛋白食品、植物肉、營養粉與調味基底中有廣泛用途,但也常伴隨豆腥味、苦味、澀感、溶解性不足或口感粗糙等問題。胺肽酶可被放在前段蛋白酶之後,針對已形成的短胜肽進行 N 端修剪,降低部分苦味胜肽的影響,並提高游離胺基酸比例。
植物蛋白加工不應只依賴單一酵素判斷成敗。原料蛋白的變性程度、抗營養因子、前處理、熱歷史與前段水解酵素選擇,都會改變胺肽酶可接觸的底物型態。從食品生物技術角度看,酵素常被用於提升食品加工效率與改善原料功能,但最終效果仍必須放回整體製程條件中理解[1]。
在調味基底開發中,蛋白水解物常提供鮮味、厚味、回甘與背景風味。若前段水解造成苦味偏高,或水解物缺乏風味深度,胺肽酶可作為後段調整工具,協助釋放更多游離胺基酸並修飾短胜肽組成。對素食肉味、發酵風味、醬汁、湯底與鹹味粉末而言,這類後段修飾有助於降低配方端對遮苦劑、香料或過量鹽分的依賴。
但風味系統的最終結果仍來自多因素交互作用。游離胺基酸可提供基礎味覺與熱反應前驅物,但熱處理條件、糖類、脂質氧化物、酵母抽出物、香辛料與鹽酸平衡都會影響終端風味。胺肽酶的功能是改變水解物的可反應與可感知組成,而不是替代配方設計。LTA4H aminopeptidase 的研究顯示,小分子與突變可改變胺肽酶底物特異性,這也從分子層級說明「底物選擇性」是胺肽酶應用表現的核心因素[7]。
營養配方常使用蛋白水解物,是因為水解可改善溶解性、降低大分子蛋白帶來的加工負擔,並形成較容易調整的胜肽與胺基酸組成。胺肽酶可用於後段增加游離胺基酸比例、降低苦味,使高蛋白飲品、粉末配方或特殊營養產品更容易達到可接受的感官表現。
需要保守區分的是,使用胺肽酶進行蛋白水解,不等同於終端產品具有特定健康功效。某些蛋白胜肽確實受到營養與功能性研究關注,但胜肽功能高度依賴序列、分子量、穩定性、消化行為與人體證據。本文將 Aminopeptidase Enzyme For Protein Hydrolysis 定位為加工用酵素工具,而非直接宣稱健康效果的功能性成分;這樣的表述更符合酵素在食品生物技術中的加工角色[1]。
膠原、明膠、肉類副產物與其他動物蛋白水解物常用於胜肽原料、調味基底或營養產品。這類原料在前段通常需要有效降低分子量,建立可溶性與穩定的水解輪廓;胺肽酶則更適合放在後段,針對苦味、游離胺基酸與末端胜肽組成進行調整。
在動物蛋白水解中,胺肽酶的實務價值通常體現在「補足前段水解的不足」。若前段內切型蛋白酶已產生大量短胜肽,但產品仍有明顯苦味或風味不平衡,胺肽酶可提供進一步修飾路徑。反之,若原料尚未充分水解,胺肽酶可作用的 N 端短胜肽有限,單獨使用時效果可能不明顯。這也呼應 aminopeptidase P 等酵素研究中對底物結構與序列限制的重視[5]。
胺肽酶需要可接觸的胜肽底物。若完整蛋白質仍維持緊密摺疊,或前段內切型水解不足,胺肽酶能處理的 N 端數量有限;若前段水解過度,則可能已形成複雜苦味胜肽群,後段修飾雖仍可能有幫助,但需要更精細控制。實務上,胺肽酶最合理的使用位置,是在前段蛋白酶已產生足夠短胜肽後,作為風味與游離胺基酸輪廓的後段調整工具。
從分子機制看,這是因為胺肽酶的活性位點必須辨識底物的 N 端。人類 aminopeptidase B 的研究顯示,特定胺基酸殘基會參與底物辨識,尤其影響其對鹼性胺基酸底物的選擇性;這種精細辨識能力說明,底物是否以合適形式呈現,會直接影響胺肽酶表現[8]。
酵素是蛋白質,會受到反應環境影響。pH 會改變酵素活性位點與底物胺基酸的電荷狀態;溫度會影響反應速率與酵素構形穩定性;鹽度可能改變蛋白質溶解性與酵素—底物互動;多酚、金屬螯合物、界面活性成分或高濃度糖鹽也可能改變水解表現。這些因素不一定使胺肽酶失效,但會讓相同添加邏輯在不同配方中產生不同結果。
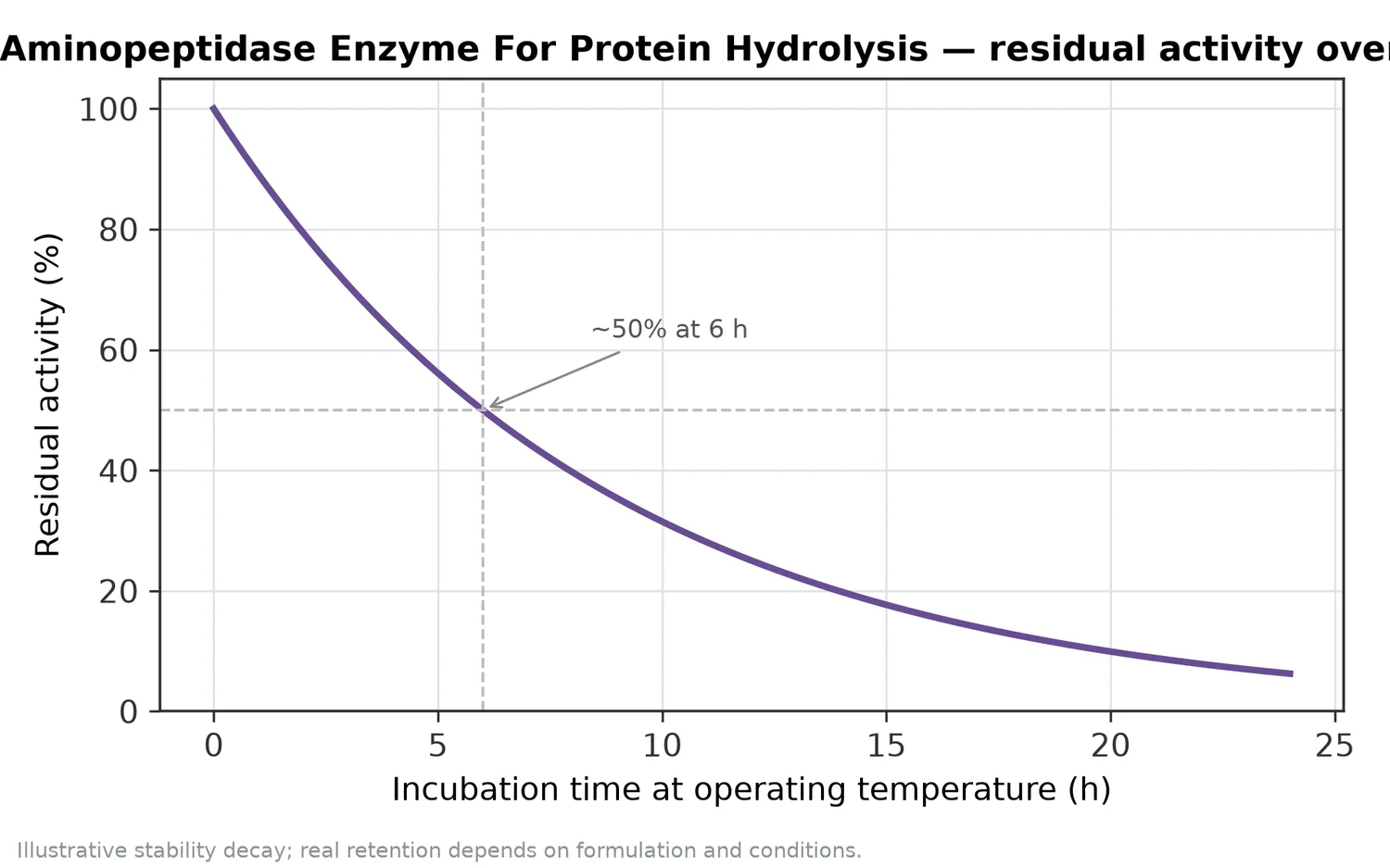
因此,Aminopeptidase Enzyme For Protein Hydrolysis 應被視為需要與原料、前處理、前段蛋白酶、反應終點及後處理共同設計的工具。酵素固定化與工業酵素應用文獻也指出,自由酵素在溫度、pH、抑制物與重複使用等情境下可能受到限制,這就是為何工業生物催化常需考量穩定性與製程相容性[9]。
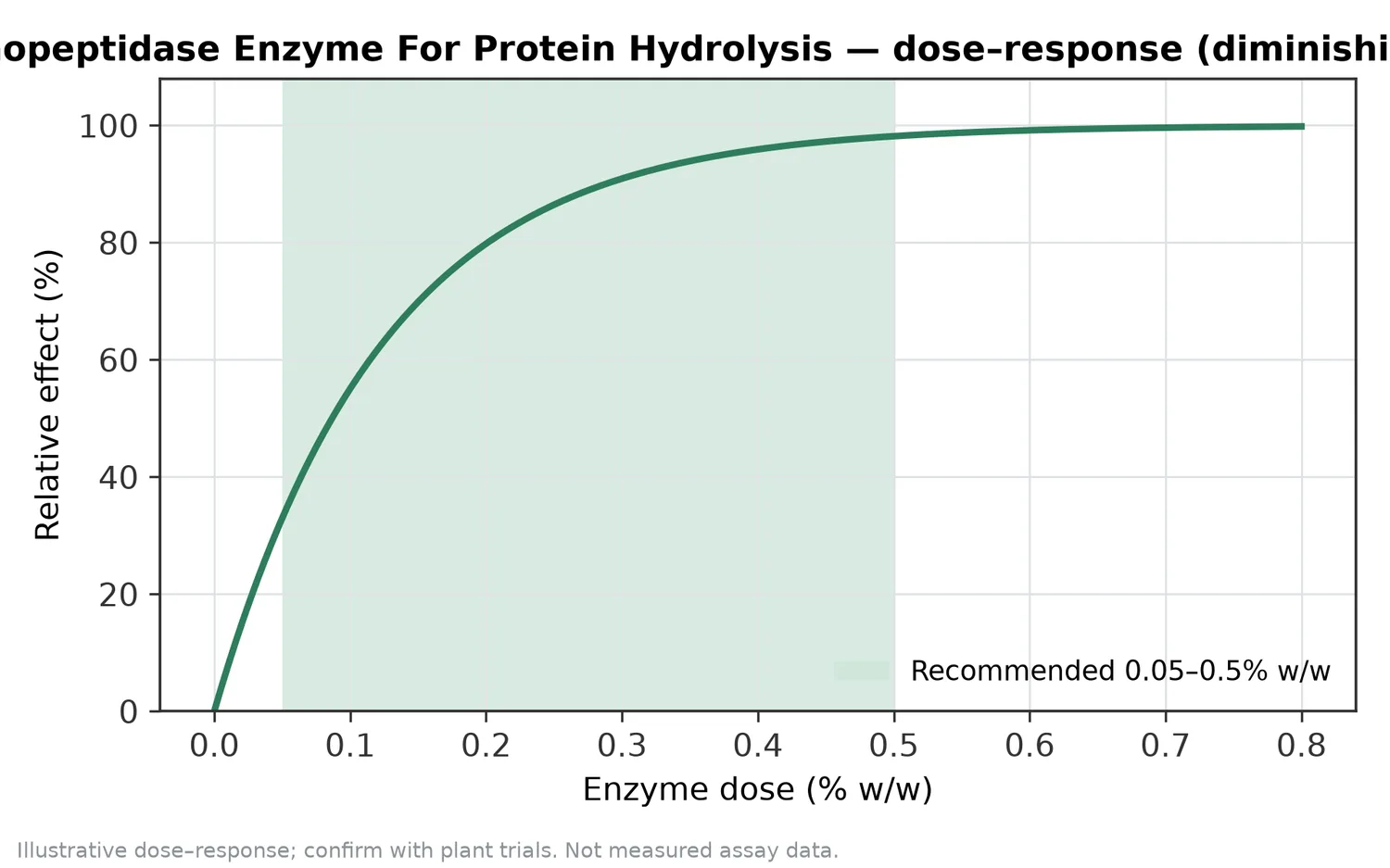
胺肽酶反應不是越久越好。若目標是降低苦味與提高游離胺基酸,反應終點需要與風味、溶解性、顏色、熱反應潛力與後段乾燥條件一起考量。過度水解可能使水解物過於單薄、鹹苦失衡,或在後續熱處理中產生過強反應風味;不足則可能保留苦味胜肽或使游離胺基酸提升有限。
後段處理也會改變水解物品質,包括加熱、濃縮、乾燥、過濾與配方混合。酵素在食品加工中常被用於提升效率與品質,但其效果必須搭配下游加工條件理解;微生物酵素在食品生物技術中的應用範圍廣泛,正是因為它們能與不同加工單元結合,而不是孤立存在[1]。
Aminopeptidase Enzyme For Protein Hydrolysis 的合理效益包括:協助降低高水解度蛋白水解物的苦味、提高游離胺基酸比例、改善調味基底的風味厚度、支援二段式蛋白水解流程,以及讓水解物組成更容易朝目標方向微調。這些效益來自胺肽酶對 N 端胜肽的逐步修剪,而不是物理遮蔽或香精補味。
需要避免的第一個誤解是把胺肽酶當作萬用去苦劑。若苦味來自原料氧化、熱裂解副產物、過度發酵、礦物鹽、植物多酚或配方中其他成分,胺肽酶只能處理與胜肽相關的部分。第二個誤解是把胺肽酶當成初段主力蛋白酶;它更適合接在內切型蛋白酶後面,處理已形成的短胜肽。第三個誤解是把使用胺肽酶與健康功效直接連結;酵素水解可改變蛋白與胜肽組成,但終端功能主張仍需獨立證據支持。
從研究角度看,不同胺肽酶在底物辨識上差異顯著。methionine aminopeptidase 研究顯示,特定突變會影響催化功能並干擾相關酵素作用,說明胺肽酶活性高度依賴其結構完整性與催化位點安排[10]。因此,在產業應用中,較準確的表述不是「胺肽酶一定能達成某個固定結果」,而是「胺肽酶可作為後段修飾工具,協助朝特定風味與胺基酸輪廓調整」。
Enzymes.bio 作為供應商,提供 Aminopeptidase Enzyme For Protein Hydrolysis 這類蛋白水解用酵素的線上購買服務,產品以 1kg 單位銷售。本文不將 Enzymes.bio 描述為製造商或實驗室,也不提供活性單位、等級、分析方法或活性定義;實際文件管理應以隨訂單提供的 CoA 與 SDS 為準。Enzymes.bio 的蛋白水解酵素分類頁可作為理解其產品應用範圍的入口。
對 B2B 使用者而言,CoA 與 SDS 的價值在於內部收貨、文件留存、安全處理與合規流程管理。酵素相關最佳實務文件強調,酵素產品需要適當處理、儲存與文件化,以維持品質並支援安全使用;這類原則適用於加工用酵素的內部管理,但不代表供應商本身執行製造或檢測工作[11]。
Aminopeptidase Enzyme For Protein Hydrolysis 最適合被理解為蛋白水解後段精修酵素。它的主要應用不是取代前段內切型蛋白酶,而是在蛋白質已被切成較短胜肽後,從 N 端逐步釋放胺基酸,協助降低部分苦味胜肽影響、提高游離胺基酸比例,並改善水解物的風味輪廓。胺肽酶結構與底物特異性研究顯示,這類酵素的效果與活性位點、S1 口袋、底物序列及反應環境密切相關,因此其應用需要放在完整製程中評估[3]。
對植物蛋白水解物、調味基底、營養配方、膠原與動物蛋白水解物而言,胺肽酶提供的是一種可控制胜肽末端與游離胺基酸組成的生物催化路徑。若使用者的目標是降低高水解度水解物苦味、提升風味厚度或建立更細緻的二段水解流程,Aminopeptidase Enzyme For Protein Hydrolysis 是值得納入製程設計的工具;但最終表現仍會受到原料、前段酵素、pH、溫度、鹽度、反應時間與後段加工共同影響。Enzymes.bio 以 1kg 單位線上供應此類產品,CoA 與 SDS 會隨訂單一併提供,供使用者進行內部文件與安全管理。
以 1 kg 單位販售,現貨供應,可立即出貨。請直接於我們的線上商店下單並付款,我們將為您處理訂單。每筆訂單皆附分析證明書與安全資料表。
購買 Aminopeptidase Enzyme For Protein Hydrolysis →依首次引用順序編號。所有來源皆為開放取用資料,並於發布時確認可連線;正文中的引用編號會連結至此。