Aminopeptidase is an exopeptidase that trims amino acids from the N-terminal end of peptides, making it especially useful after broader proteases have already broken proteins into smaller fragments. In protein hydrolysis, this “finishing” action can increase free amino acids, shorten residual peptides, and help reduce bitterness or improve savory flavor development in protein-rich food, fermentation, nutrition, and feed applications. Enzymes.bio supplies Aminopeptidase directly online by the 1 kg unit; orders are paid online, processed, and shipped with a Certificate of Analysis and Safety Data Sheet included.
The central aminopeptidase function is directional peptide trimming. Proteins and peptides are chains of amino acids with two chemically different ends: the N-terminus, which carries the free amino group, and the C-terminus, which carries the free carboxyl group. Aminopeptidases act at the N-terminal end, releasing one amino acid at a time when the peptide sequence, enzyme structure, and process environment are compatible. Structural and biochemical work on aminopeptidases, including the LapB aminopeptidase from Legionella pneumophila, supports this view of aminopeptidases as enzymes specialized for N-terminal peptide-bond hydrolysis rather than random internal cutting [1].
That makes aminopeptidase different from the broad proteases often used at the start of protein hydrolysis. Endoproteases open up a folded or aggregated protein by cutting internal peptide bonds, creating a mixture of shorter peptides. Aminopeptidase then works on the exposed N-termini of those peptides. In practice, the result is not simply “more hydrolysis” in a generic sense: the peptide population shifts toward shorter chains, and the free amino acid fraction increases because residues are released sequentially from peptide ends.
This directional mechanism is why aminopeptidase is often described as a polishing or finishing enzyme in protein hydrolysate systems. A hydrolysate produced with only an endoprotease may still contain medium-length peptides that taste bitter, bind water differently, or behave unpredictably in heat processing. Adding aminopeptidase activity can change the hydrolysate profile by clipping exposed N-terminal residues and reducing the concentration of intact bitter peptides. In a chicken breast meat hydrolysis study using leucine aminopeptidase produced by Aspergillus oryzae LL1, the enzyme was applied specifically to improve hydrolysis of a protein-rich animal substrate, illustrating the industrial relevance of leucine aminopeptidase in meat-protein processing [2].
At the molecular level, aminopeptidase changes a protein hydrolysate in three connected ways. First, it increases the number of free amino acids by cleaving the first peptide bond after the N-terminal residue. Second, it shortens peptides stepwise, which can alter solubility, taste, and reactivity. Third, it can expose a new N-terminus after each cleavage, allowing further trimming if the next residue fits the enzyme’s active site.
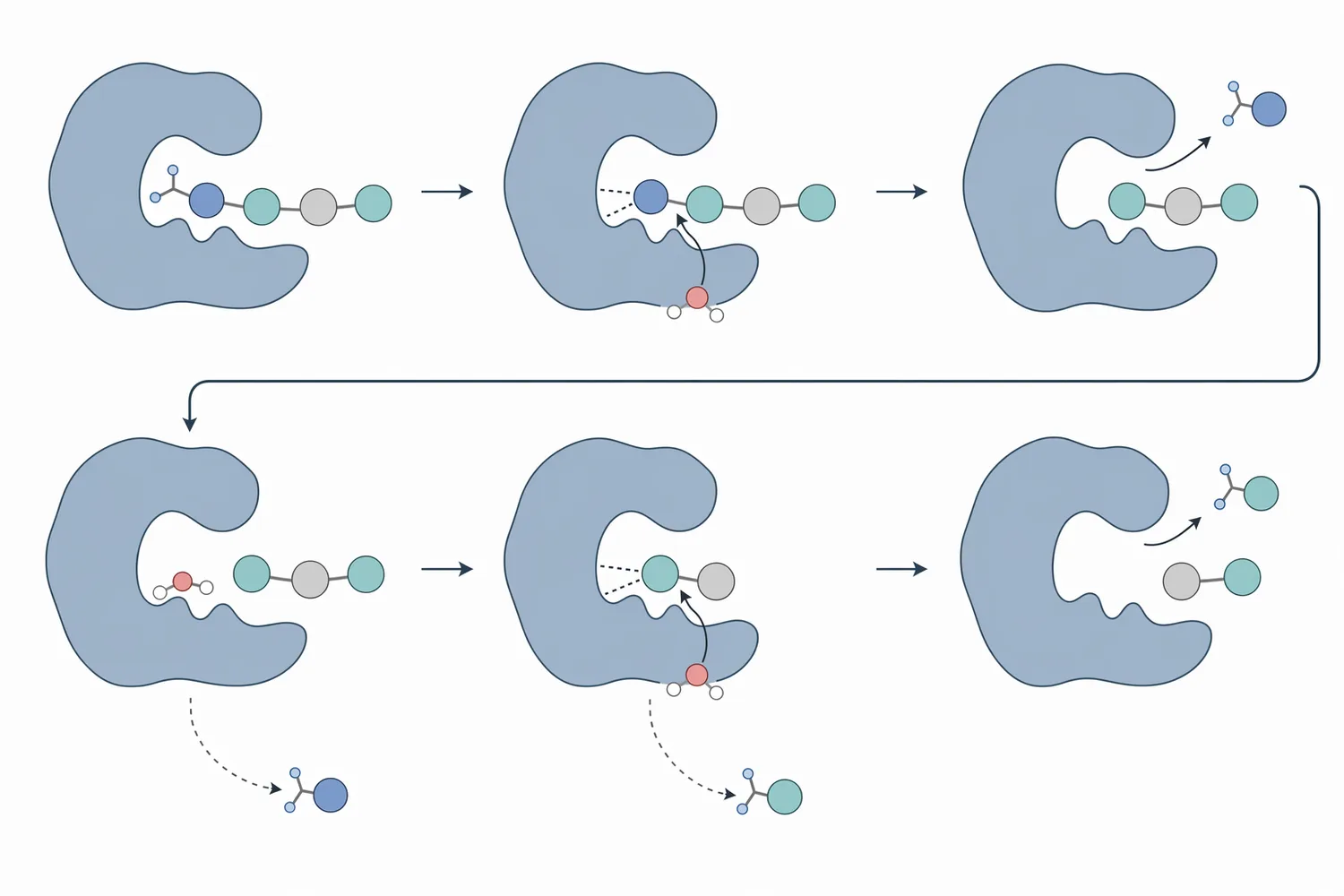
Many aminopeptidases are metalloenzymes. In those enzymes, one or more metal ions help polarize and activate a water molecule so that it can attack the carbonyl carbon of the peptide bond. The peptide bond then breaks, releasing the N-terminal amino acid and leaving behind a peptide that is one residue shorter. A mechanistic investigation of Aeromonas proteolytica aminopeptidase using quantum mechanical/molecular mechanical modeling examined this metal-assisted mode of action, showing why active-site geometry and metal coordination are central to peptide-bond cleavage [3].
This mechanism also explains why aminopeptidase is not interchangeable with every other protease. If the substrate is a large intact protein with few accessible ends, aminopeptidase has limited access until an endoprotease or fermentation process generates peptides. If the substrate is already a peptide-rich hydrolysate, aminopeptidase has many more N-terminal sites to work on. This is why aminopeptidase and dipeptidase activities are often discussed together in digestion and fermentation contexts: aminopeptidases remove residues from longer peptides, while dipeptidases complete the breakdown of dipeptides into individual amino acids.
Amino acid sequence matters as well. Terms such as leucine aminopeptidase, leucyl aminopeptidase, alanyl aminopeptidase, aminopeptidase B, and leucyl-cystinyl aminopeptidase refer to enzyme types or naming traditions linked to substrate preference, biological origin, or historical characterization. They should not be read as identical products. For example, leucine aminopeptidase is commonly associated with N-terminal leucine-containing substrates, while methionine aminopeptidase has the specialized biological role of removing the initiator methionine from newly synthesized proteins. Reviews of methionine aminopeptidase 2 describe this N-terminal methionine removal as a conserved protein-maturation step, which is why a methionine aminopeptidase 2 inhibitor is studied in therapeutic research rather than as a food-processing enzyme [4].
The practical distinction between aminopeptidase vs trypsin is the location of the cut. Trypsin is an endoprotease: it cuts inside peptide chains at specific internal sites. Aminopeptidase is an exopeptidase: it trims from the N-terminal end. When buyers compare trypsin vs aminopeptidase, the useful question is not which enzyme is “stronger,” but what kind of peptide profile is needed. Internal cleavage creates more peptide ends; N-terminal trimming converts those peptide ends into free amino acids and shorter fragments.
| Enzyme type | Main cutting position | What changes in the substrate | Typical processing role |
|---|---|---|---|
| Aminopeptidase | N-terminal end of peptides | Releases one N-terminal amino acid at a time; shortens peptides sequentially | Finishing hydrolysis, debittering support, free amino acid release |
| Trypsin-type endoprotease | Internal peptide bonds | Opens proteins and produces multiple peptide fragments | Primary protein breakdown and peptide generation |
| Carboxypeptidase | C-terminal end of peptides | Releases amino acids from the opposite end of the peptide chain | Complementary exopeptidase action |
| Dipeptidase | Dipeptides | Splits two-residue peptides into free amino acids | Final digestion or complete peptide breakdown |
Aminopeptidase and carboxypeptidase can therefore be complementary rather than competitive: one works from the amino end, the other from the carboxyl end. Aminopeptidase and dipeptidase are also complementary, because aminopeptidase can reduce longer peptides to shorter peptides, while dipeptidases act when the remaining substrate is already a two-amino-acid unit. Heavy-oxygen exchange studies on proteolytic enzymes show how different proteases can be distinguished mechanistically by how they cleave peptide bonds and exchange oxygen during catalysis, reinforcing that “protease” is a broad category rather than one uniform mode of action [5].
Aminopeptidases are widely distributed in microorganisms, animals, and human tissues. When people ask where aminopeptidase is found, the answer depends on the enzyme family: aminopeptidase N is a membrane-associated enzyme in many animal systems, methionine aminopeptidase is part of intracellular protein maturation, microbial leucyl aminopeptidases occur in fungi and bacteria, and specialized aminopeptidase complexes are found even in extremophiles. A TET-like aminopeptidase complex from the hyperthermophilic archaeon Pyrococcus horikoshii illustrates how aminopeptidase function is conserved even in organisms adapted to very high-temperature environments [6].
When the question is where aminopeptidase is produced for food-enzyme use, microbial sources are especially important. Food-enzyme safety evaluations have assessed leucyl aminopeptidase from non-genetically modified Aspergillus strains, including Aspergillus oryzae strain NZYM-EX and another Aspergillus strain designated AE-MB. These evaluations are relevant because Aspergillus species are widely used as enzyme sources in food-processing contexts, and they show that leucyl aminopeptidase is a recognized food-enzyme category rather than only a research enzyme [7].
The biological breadth of aminopeptidases also explains why similar names can appear in very different fields. Aminopeptidase N has been studied in reproductive biology, where elevated aminopeptidase N was associated with effects on sperm motility and early embryo development in an animal study [8]. Insulin-regulated aminopeptidase has been reviewed in relation to angiotensin IV signaling [9]. These biomedical examples are not claims for industrial food use; they show that aminopeptidase activity is biologically important and that enzyme naming often reflects tissue location or physiological role.
Leucine aminopeptidase, also called leucyl aminopeptidase in many regulatory and technical contexts, is one of the most relevant aminopeptidase types for protein hydrolysis and flavor development. Its value comes from its ability to act on peptides with suitable N-terminal residues and release amino acids that can directly affect taste or serve as flavor precursors. In protein-rich food matrices, this can mean fewer residual bitter peptides and more soluble nitrogen in forms that contribute to savory, brothy, or fermented notes.

The Aspergillus oryzae LL1 chicken breast meat hydrolysis study is a useful application example because it connects leucine aminopeptidase to a real protein substrate rather than a purified laboratory peptide. Chicken breast meat contains myofibrillar and sarcoplasmic proteins that form complex peptide mixtures during hydrolysis. Leucine aminopeptidase can act after larger proteins have been opened into accessible peptides, releasing N-terminal amino acids and changing the peptide distribution in a way that is difficult for an endoprotease alone to achieve [2].
Food-enzyme safety evaluations of leucyl aminopeptidase from non-genetically modified Aspergillus strains further support the relevance of this enzyme type for food-processing contexts. EFSA’s evaluation of leucyl aminopeptidase from Aspergillus sp. strain AE-MB considered its intended use as a food enzyme, while a separate evaluation covered leucyl aminopeptidase from Aspergillus oryzae strain NZYM-EX [10]. These sources do not mean every aminopeptidase product is the same; they show that leucyl aminopeptidase is an established enzyme category with documented food-enzyme assessments.
Bitterness in protein hydrolysates is often caused by peptides rather than by intact proteins. When hydrolysis exposes hydrophobic amino acid residues inside a peptide, those peptides can interact with bitterness receptors and create an undesirable aftertaste. Aminopeptidase helps because it does not merely chop randomly; it progressively removes N-terminal residues from peptides that are accessible to the enzyme. If bitter character is associated with a peptide’s exposed N-terminal sequence or with the peptide’s length and hydrophobicity, aminopeptidase action can reduce the intensity of that sensory defect.
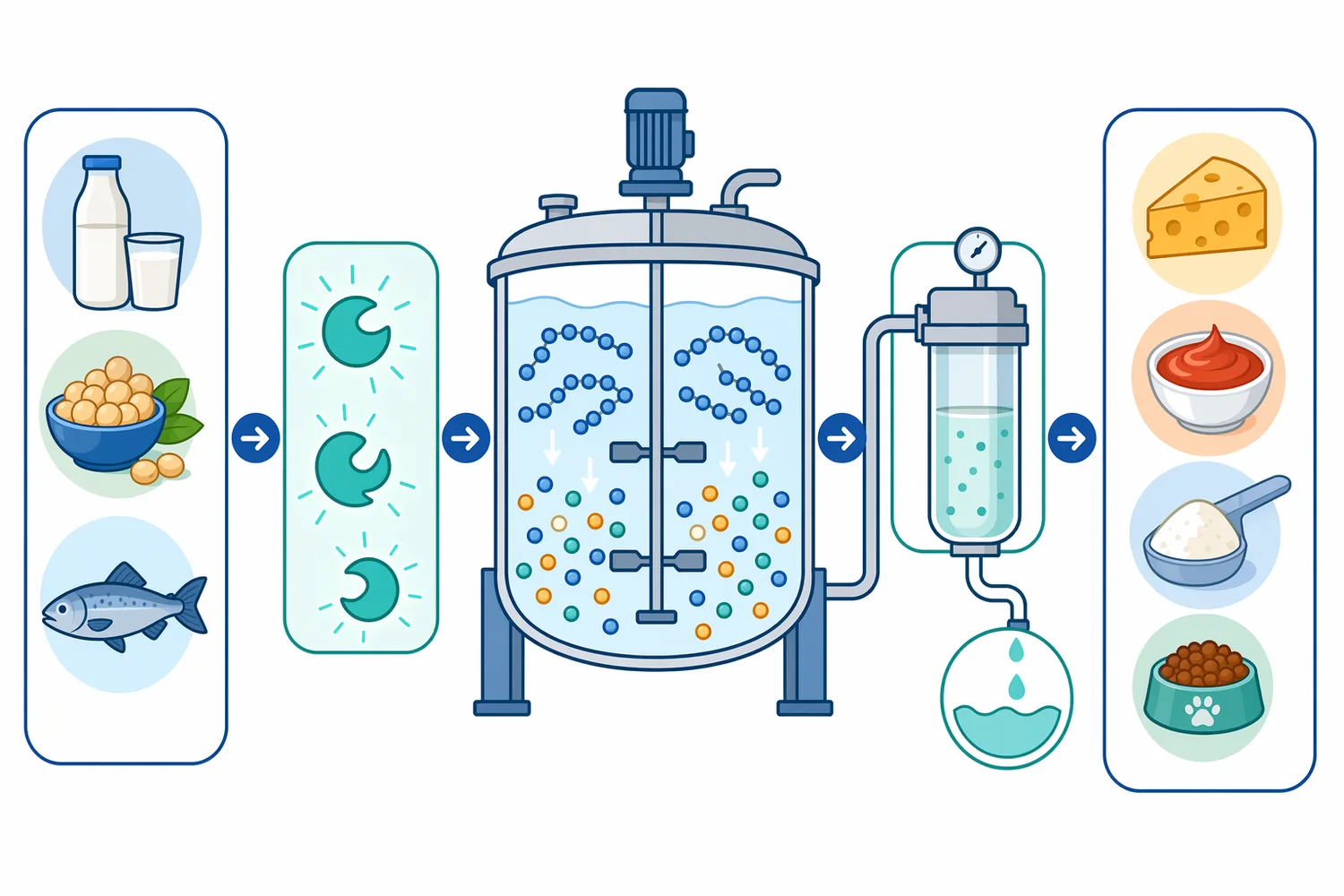
This is especially relevant for hydrolysates from meat, fish, dairy, plant proteins, oilseed meals, and by-products where nutritional value is high but taste is limiting. In such systems, the enzyme’s practical contribution is a shift in the peptide and amino acid balance. Some peptides disappear or become shorter; some free amino acids increase; and the hydrolysate may become easier to formulate into seasonings, nutritional blends, savory bases, or feed ingredients. The chicken breast hydrolysis work with A. oryzae leucine aminopeptidase demonstrates this type of use case in an animal-protein matrix [2].
A related example comes from bi-enzyme hydrolysis of Manchurian walnut dreg protein. Walnut dreg is a protein-containing by-product, and the study examined how a two-enzyme hydrolysis approach changed the properties and composition of the resulting hydrolysates. While the exact enzyme system and target properties are specific to that study, the broader principle is directly relevant: combining enzymes can produce hydrolysates with different composition and functional characteristics than a single-step process [11].
In fermented and ripened foods, aminopeptidase contributes by releasing amino acids from peptides generated during protein breakdown. Those amino acids can taste sweet, bitter, umami-like, sulfurous, or neutral depending on the residue and food matrix. They can also become precursors for later reactions: transamination, decarboxylation, Strecker degradation, sulfur-compound formation, and Maillard-type reactions all depend on the amino acid pool available during processing.
Cheese is a clear example. During ripening, caseins are progressively degraded into peptides and amino acids. The sensory profile of cheese depends not only on how much protein is hydrolyzed but on which peptides remain and which amino acids are released. A study on high-pressure processing of cheese made from unpasteurized milk examined microbiology, proteolysis, biogenic amines, and flavor, showing how proteolytic changes are intertwined with ripening chemistry and sensory development [12].
Aminopeptidase is also relevant to fermented seafood, soy-type condiments, protein seasonings, and other savory systems where microbes and enzymes convert proteins into soluble nitrogen compounds. In Thai traditional fermented shrimp paste, microbial diversity changes during processing, and such microbial succession is part of the biochemical environment that drives protein breakdown and flavor formation [13]. Commercial aminopeptidase use applies the same biochemical logic in a more directed way: increase N-terminal trimming of peptides to support controlled flavor development.
Tea processing is not a protein hydrolysis application in the same way as cheese or meat, but it shows how microbial and enzymatic functions can shift during food processing. Research on black tea processing reported dynamic changes in bacterial community diversity and function, highlighting that food-processing matrices are active biochemical systems rather than inert substrates [14]. For aminopeptidase users, the lesson is practical: enzyme performance must be understood in the context of the whole matrix, including pH, competing enzymes, salt, heat history, and substrate accessibility.
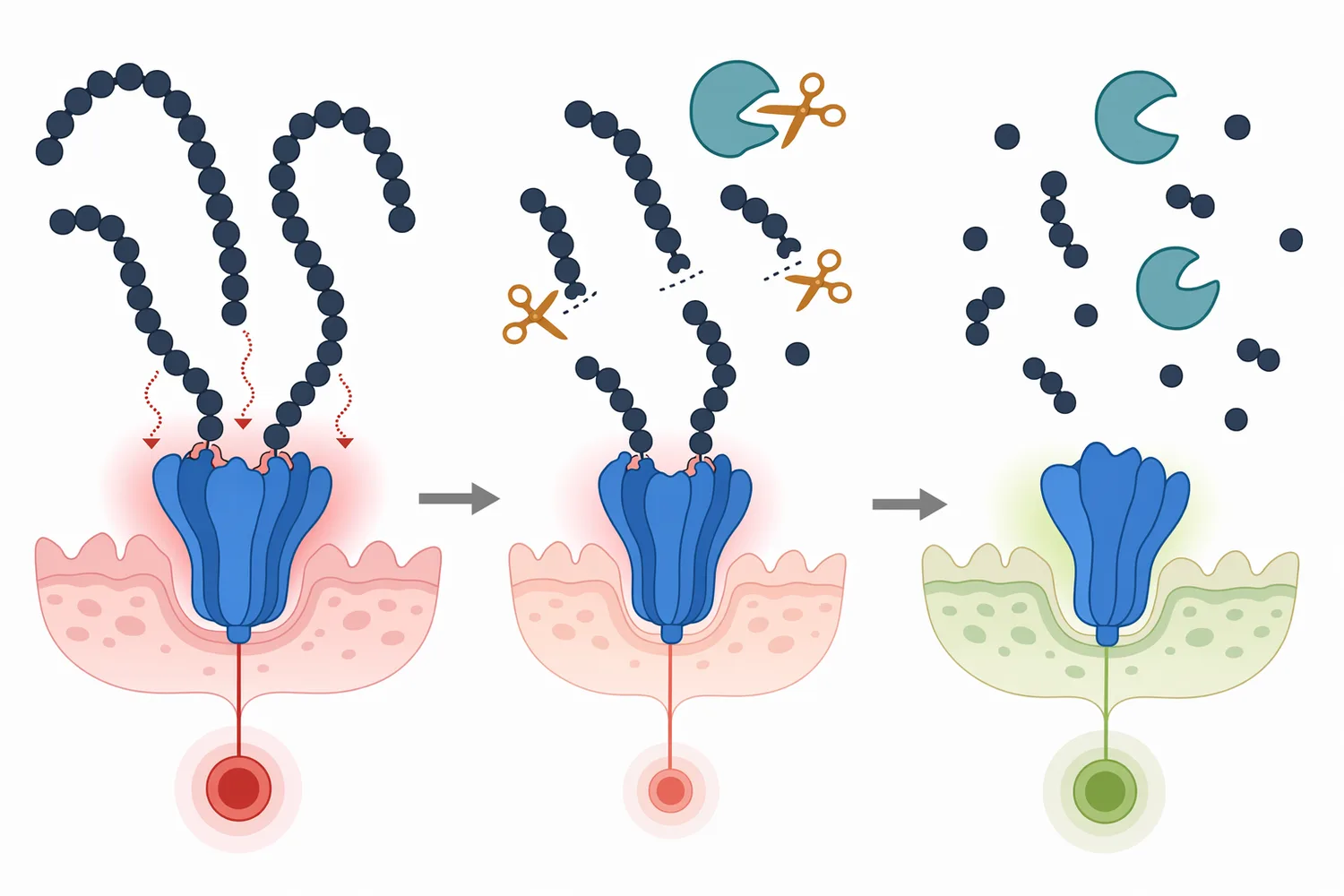
Aminopeptidase has a natural role in digestion because proteins must ultimately be converted into absorbable amino acids and small peptides. In animal nutrition, this makes aminopeptidase relevant where protein sources are difficult to digest, where hydrolysates are used for young animals, or where by-products are upgraded into more usable protein ingredients. The enzyme’s role is not to add protein; it changes the form in which protein nitrogen appears.
A zebrafish nutrition study examining the effects of protein and fiber content on gut structure and function used digestive-function measurements, including aminopeptidase activity, as part of the evaluation of how diet composition affects intestinal physiology [15]. This kind of evidence supports the biological importance of brush-border and intestinal peptidase activity in protein utilization. Industrial aminopeptidase applications are not identical to in vivo digestion, but they use the same core chemistry: peptide-end hydrolysis to increase the pool of smaller peptides and free amino acids.
For feed or nutrition applications, aminopeptidase is most relevant where a protein substrate has already been disrupted by heat, grinding, fermentation, or other proteases. If proteins remain insoluble, highly crosslinked, or physically inaccessible, aminopeptidase has fewer N-terminal peptide ends to attack. When prior processing exposes peptides, aminopeptidase can help finish the hydrolysis profile and may improve the consistency of the resulting ingredient.
Not every aminopeptidase used in research is intended for food or industrial hydrolysis. Methionine aminopeptidase removes the initiator methionine from newly synthesized proteins, a process required for normal protein maturation in cells. Methionine aminopeptidase 2, often abbreviated MetAP2, has become a biomedical target because blocking it can affect pathways connected with cancer, obesity, and immune regulation. That is why terms such as methionine aminopeptidase 2 inhibitor appear frequently in pharmaceutical literature [4].
Newer reviews of MetAP2 as a therapeutic target discuss structural insights, mechanistic roles, and inhibitor development for obesity and type 2 diabetes research. These studies are important scientifically, but they should not be confused with the use of aminopeptidase as a processing enzyme for food or protein ingredients. A methionine aminopeptidase inhibitor is designed to suppress a biological enzyme target; a processing aminopeptidase is used to hydrolyze peptide substrates outside that therapeutic context [16].
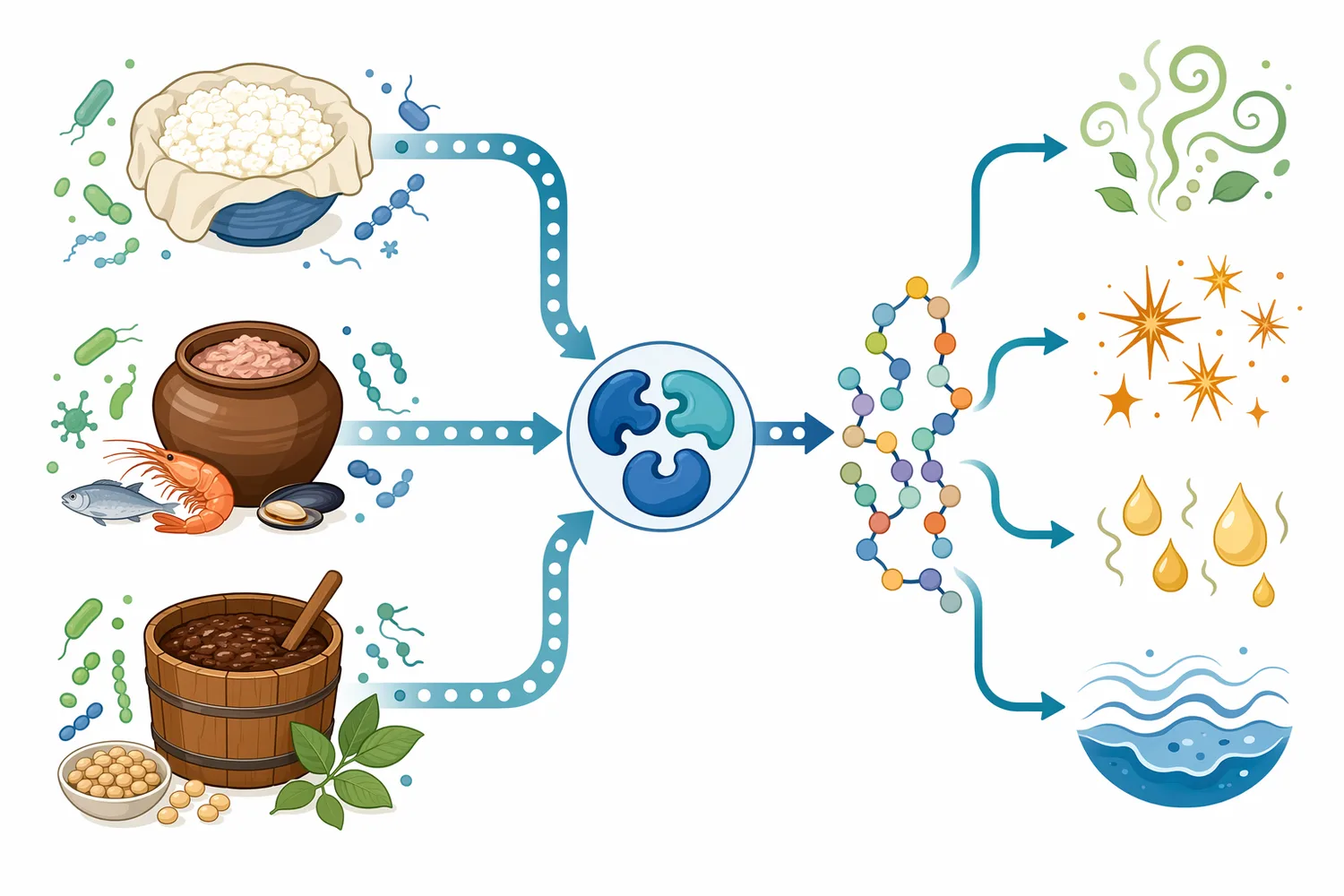
Aminopeptidase N is another specialized term. In biomedical literature, aminopeptidase N inhibitors may be studied because aminopeptidase N participates in cell-surface peptide metabolism and disease-related processes. Dual aminopeptidase inhibition has also been investigated for cross-species antimalarial activity, showing that aminopeptidases can be drug targets in pathogens as well as useful processing enzymes in industry [17]. Again, the shared word “aminopeptidase” refers to a biochemical cleavage function, not a single product or application.
Aminopeptidase performance depends on the enzyme type and the processing matrix. The most important practical point is substrate accessibility: aminopeptidase needs exposed N-termini. Large proteins may need prior unfolding, fermentation, or endoprotease treatment before aminopeptidase can contribute meaningfully. Once peptides are available, pH, temperature, salt, metal ions, and inhibitors can influence the enzyme’s active-site structure and substrate binding.
High-pressure processing provides a useful illustration of how physical treatment can alter enzyme structure and function. A purified X-prolyl dipeptidyl aminopeptidase from Streptococcus thermophilus was studied under high pressure to evaluate structural modifications and enzymatic activity. The study is enzyme-specific, but it demonstrates a general principle: processing conditions can change enzyme conformation and therefore catalytic performance [18].
Matrix composition matters just as much as the enzyme itself. In a dense meat slurry, a fermented dairy curd, a plant-protein hydrolysate, or a high-salt condiment mash, peptide diffusion, protein solubility, water availability, and competing reactions are different. Aminopeptidase may show strong activity on soluble peptides but much less effect on aggregated or insoluble protein fractions. This is why aminopeptidase is often most effective when integrated into a process that already creates soluble peptide substrates.
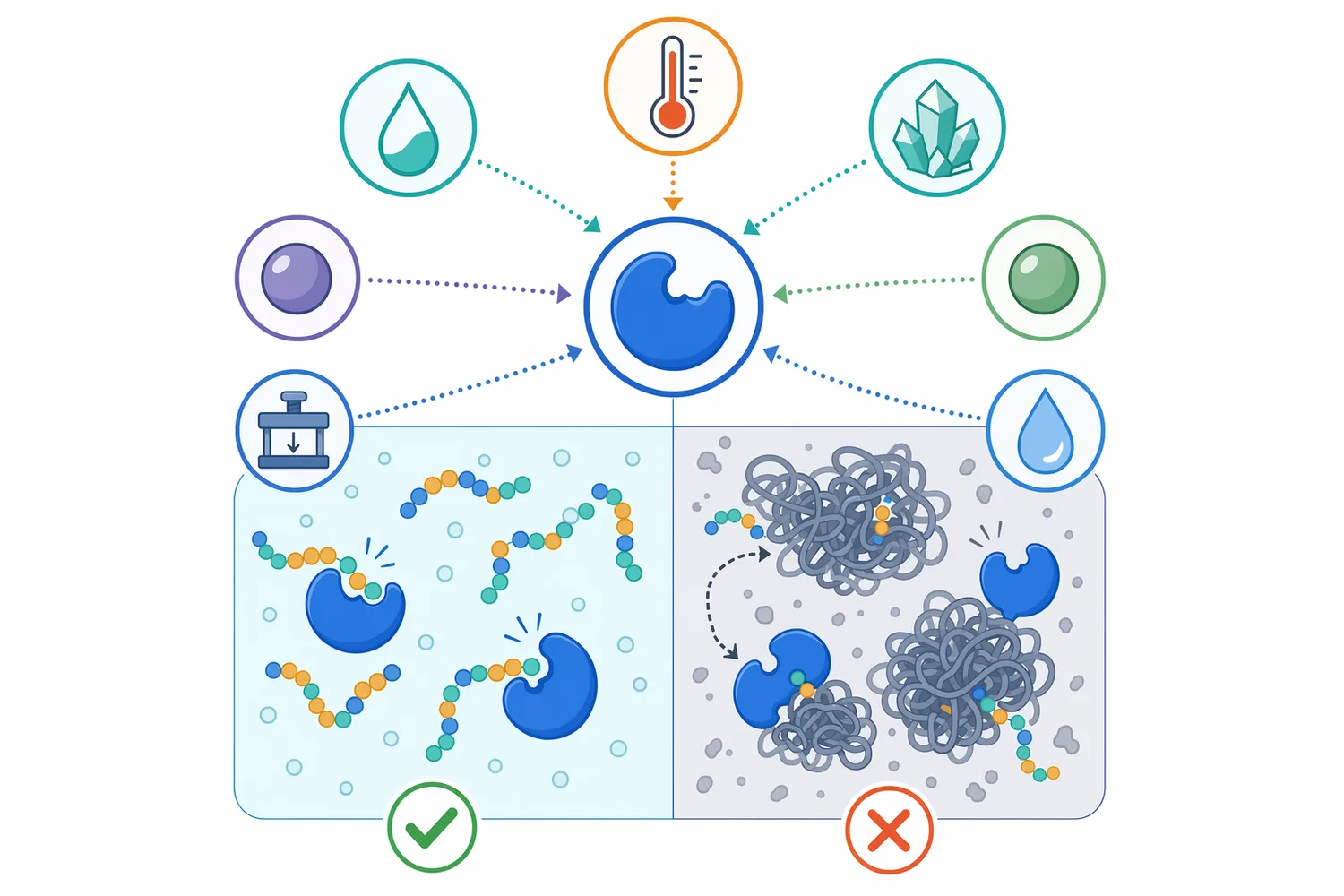
Aminopeptidase is used to refine protein hydrolysates by increasing free amino acids and reducing residual peptide length. This is valuable in nutrition powders, savory bases, meat hydrolysates, plant-protein ingredients, collagen or gelatin hydrolysates, and specialty feed ingredients. The main substrate change is a measurable shift away from longer peptides and toward shorter peptides and amino acids, which can influence taste, solubility, and downstream processing behavior.
The chicken breast hydrolysis study with leucine aminopeptidase from Aspergillus oryzae LL1 is directly relevant to meat-protein applications. Poultry muscle contains structured proteins that require controlled hydrolysis to become flavor-active or nutritionally modified ingredients. Leucine aminopeptidase contributes after peptide fragments are available, trimming N-terminal amino acids and changing the hydrolysate composition [2].
In cheese and fermented dairy, aminopeptidase supports the conversion of casein-derived peptides into amino acids. This can help ripening because amino acids are central to cheese flavor chemistry. Proteolysis is also connected to biogenic amine formation, texture, and sensory development, as shown in the high-pressure cheese study that evaluated proteolysis, biogenic amines, and flavor together [12].
Plant-protein by-products often contain valuable protein but may require hydrolysis to improve solubility, taste, or usability. The Manchurian walnut dreg protein study shows how bi-enzyme hydrolysis can alter hydrolysate composition and properties, supporting the broader case for enzyme combinations in by-product valorization [11]. Aminopeptidase can be part of this strategy when the goal is peptide trimming and amino acid release.
In fermented condiments, aminopeptidase can help generate free amino acids and small peptides that contribute to savory complexity. The enzyme is most relevant in protein-rich systems where other proteolytic steps have already generated peptide substrates. Traditional fermentations such as shrimp paste demonstrate how microbial communities and protein breakdown evolve together during processing [13].
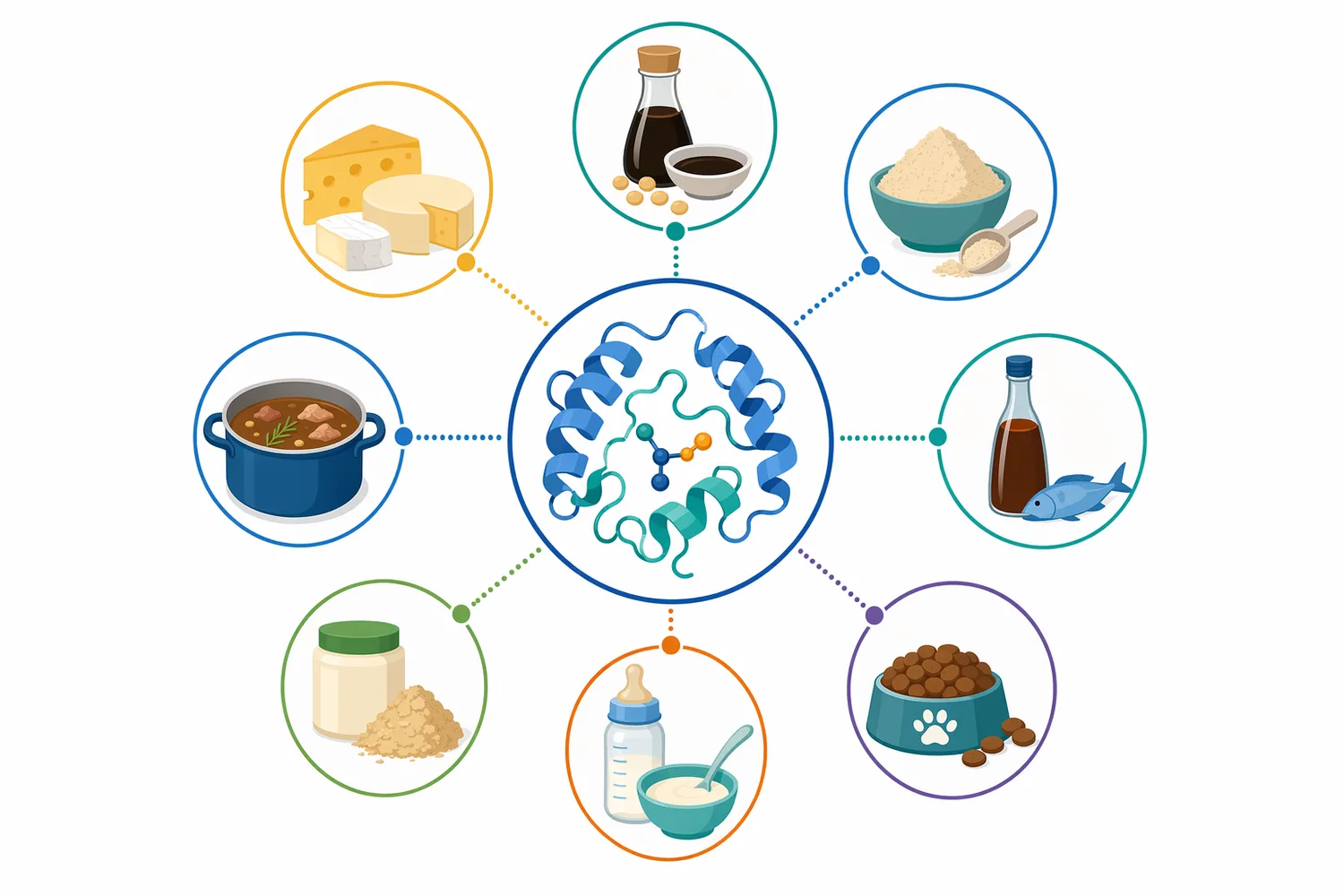
Enzymes.bio supplies Aminopeptidase as a professional enzyme product sold directly online by the 1 kg unit. The purchase process is simple: the buyer places the order online, pays online, and the order is processed and shipped. A Certificate of Analysis and Safety Data Sheet are included with the order.
This product is intended for buyers who already understand their application context and need an aminopeptidase enzyme for protein hydrolysis, peptide trimming, debittering support, flavor development, or related process work. The key scientific point is straightforward: aminopeptidase works from the N-terminal end of peptides, so its best value appears when peptide substrates are already available and the process benefits from additional amino acid release.
Aminopeptidase should be viewed as a targeted exopeptidase rather than a universal substitute for endoproteases. It complements enzymes that open proteins internally, and it can help refine the final hydrolysate profile by converting exposed peptide ends into free amino acids. For protein-rich food, fermentation, nutrition, and feed applications, that N-terminal trimming mechanism is the reason aminopeptidase remains a practical enzyme choice.
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Aminopeptidase →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.