Asparaginase, częściej opisywana w literaturze jako L-asparaginase, to enzym katalizujący hydrolizę L-asparaginy do L-asparaginianu i amoniaku. Jej praktyczna wartość polega na selektywnym zmniejszaniu dostępności L-asparaginy w układach biologicznych, technologicznych i badawczych, zwłaszcza tam, gdzie ten aminokwas jest niepożądanym substratem dalszych reakcji [1]. Enzymes.bio dostarcza asparaginase jako produkt B2B dostępny online w jednostkach 1 kg; CoA i SDS są dostarczane wraz z zamówieniem.
Asparaginase to nazwa funkcjonalna enzymu, którego głównym substratem jest L-asparagina — naturalny aminokwas obecny w organizmach żywych, surowcach roślinnych, pożywkach, hydrolizatach białkowych i wielu matrycach biologicznych. W ujęciu biochemicznym L-asparaginase należy do enzymów hydrolitycznych: wykorzystuje cząsteczkę wody do rozszczepienia grupy amidowej L-asparaginy, prowadząc do powstania L-asparaginianu oraz amoniaku [1].
Reakcję można zapisać stechiometrycznie w prosty sposób:
1 mol L-asparaginy + 1 mol H2O → 1 mol L-asparaginianu + 1 mol NH3/NH4+
Dla użytkownika procesowego kluczowe są dwie konsekwencje tej reakcji. Po pierwsze, enzym obniża stężenie wolnej L-asparaginy, czyli usuwa substrat dostępny dla kolejnych reakcji chemicznych lub metabolicznych. Po drugie, wprowadza do układu produkty o innym znaczeniu technologicznym: L-asparaginian oraz amoniak, który w zależności od pH występuje częściowo jako jon amonowy [1].
W praktyce nazwy „asparaginase” i „L-asparaginase” są często stosowane równolegle, ale druga jest bardziej precyzyjna, ponieważ wskazuje na naturalny izomer aminokwasu. To ważne w dokumentacji technicznej: działanie enzymu nie polega na ogólnym „rozbijaniu aminokwasów”, lecz na ukierunkowanej reakcji wobec L-asparaginy. Z tego powodu asparaginase jest narzędziem do problemów związanych z konkretnym aminokwasem, a nie uniwersalnym dodatkiem korygującym skład wszystkich frakcji azotowych [2].
Najprostsze wyjaśnienie mechanizmu brzmi: L-asparaginase rozpoznaje L-asparaginę, wiąże ją w centrum aktywnym i katalizuje hydrolizę jej grupy amidowej. Nie jest proteazą i nie „tnie” białek na peptydy; działa na wolny aminokwas lub dostępną formę substratu w środowisku reakcji. Ta różnica ma znaczenie, ponieważ zastosowanie enzymu ma sens przede wszystkim wtedy, gdy wolna L-asparagina jest dostępna dla centrum aktywnego [1].
Na poziomie strukturalnym wiele bakteryjnych L-asparaginaz opisuje się jako białka oligomeryczne, często funkcjonujące jako tetramery zbudowane z czterech podjednostek. W takich enzymach miejsca aktywne są formowane przez ułożenie elementów sąsiednich podjednostek, dlatego zachowanie prawidłowej struktury przestrzennej jest bezpośrednio związane z aktywnością katalityczną [1].
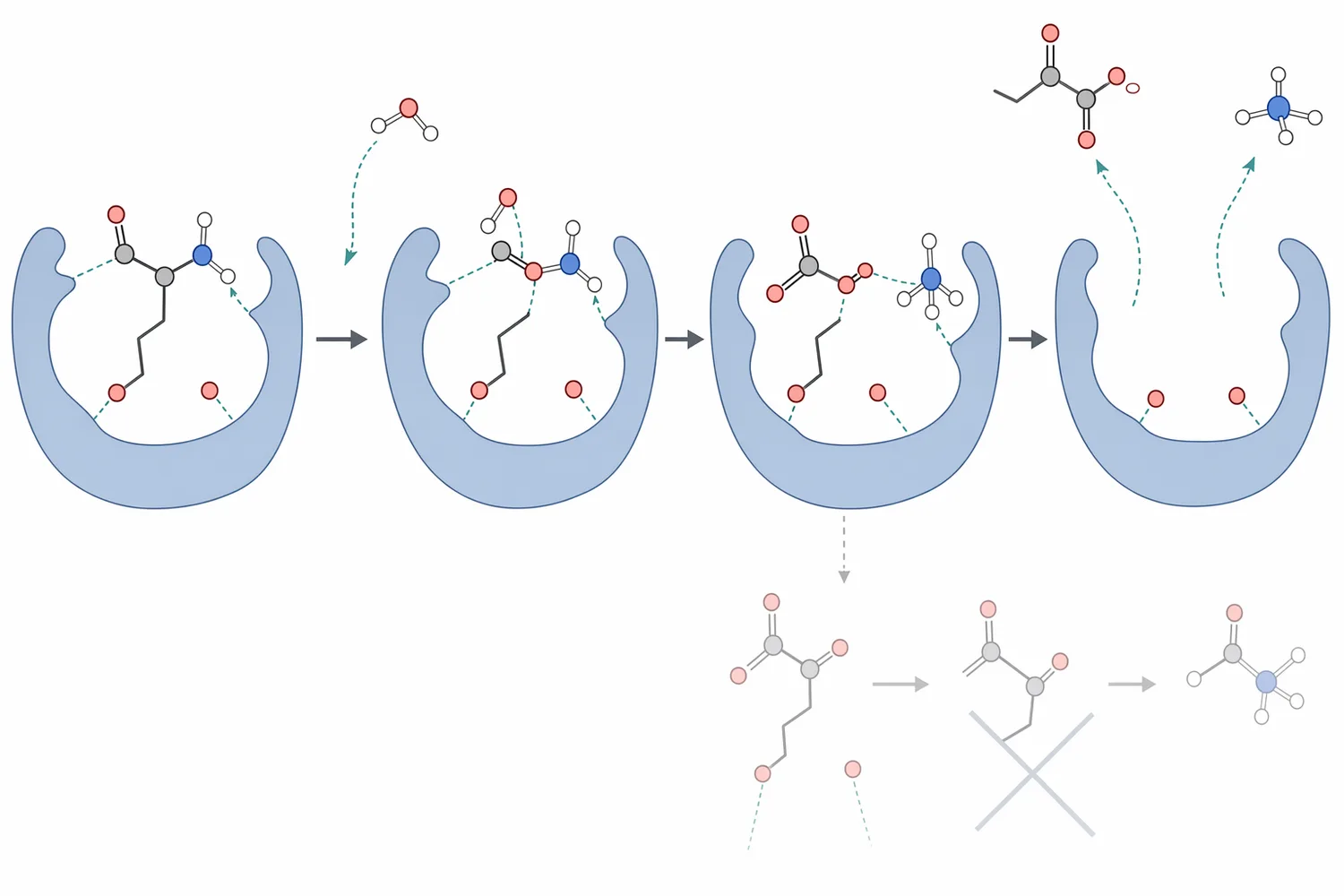
Dla części L-asparaginaz opisano również aktywność uboczną wobec L-glutaminy, czyli zdolność do hydrolizy innego aminokwasu amidowego. Z technologicznego punktu widzenia jest to istotna cecha różnicująca preparaty i warianty enzymatyczne: im bardziej proces wymaga selektywnego usuwania L-asparaginy, tym większe znaczenie ma profil substratowy enzymu, a nie sama ogólna etykieta „asparaginase” [1].
W literaturze występują różne klasy i pochodzenia L-asparaginaz. Oprócz klasycznych enzymów bakteryjnych opisywane są również enzymy typu III, takie jak EcAIII z Escherichia coli, zaliczane do hydrolaz N-końcowego nukleofila i dojrzewające przez autoproteolityczne cięcie prekursora. Takie różnice strukturalne pokazują, że ta sama funkcja — hydroliza L-asparaginy — może być realizowana przez białka o odmiennych mechanizmach dojrzewania, stabilności i profilu aktywności [3].
W procesach B2B asparaginase jest interesująca nie dlatego, że jest „enzymem specjalistycznym” samym w sobie, lecz dlatego, że L-asparagina może pełnić rolę niepożądanego prekursora. Jeśli ten aminokwas bierze udział w dalszej reakcji, która obniża jakość produktu lub utrudnia kontrolę procesu, jego wcześniejsze enzymatyczne przekształcenie może zmienić przebieg całego układu [2].
Najbardziej rozpoznawalnym przykładem jest technologia żywności, gdzie L-asparagina jest jednym z kluczowych prekursorów akrylamidu powstającego podczas intensywnej obróbki cieplnej produktów zawierających cukry redukujące. L-asparaginase działa przed tym etapem: konwertuje L-asparaginę do L-asparaginianu, który nie uczestniczy w tym samym szlaku tworzenia akrylamidu, dzięki czemu zmniejsza dostępność krytycznego substratu [2].
Ta logika jest bardziej ogólna niż sama redukcja akrylamidu. W dowolnej matrycy, w której L-asparagina jest punktem startowym reakcji ubocznej, narzędziem kontrolnym może być jej enzymatyczna deamidacja. Zastosowanie asparaginase ma więc sens wtedy, gdy zidentyfikowano związek przyczynowy: obecność L-asparaginy → niepożądany produkt, niepożądany profil reakcji lub zakłócenie interpretacji doświadczenia [1].
Ważne jest jednak odróżnienie reakcji enzymatycznej od wyniku technologicznego. Sama hydroliza L-asparaginy jest dobrze opisana, ale efekt w konkretnej matrycy zależy od tego, czy enzym ma kontakt z substratem, czy środowisko zawiera wystarczającą ilość wody, czy pH i temperatura nie dezaktywują białka oraz czy składniki receptury nie ograniczają dostępności L-asparaginy [2].
| Obszar zastosowania | Problem procesowy lub badawczy | Jak pomaga L-asparaginase | Najważniejsze ograniczenie interpretacji |
|---|---|---|---|
| Technologia żywności | Wolna L-asparagina jako prekursor reakcji podczas obróbki cieplnej | Deamiduje L-asparaginę przed etapem wysokotemperaturowym | Skuteczność zależy od wilgotności, czasu kontaktu i dostępności substratu |
| Biotechnologia i fermentacja | Potrzeba modyfikacji profilu aminokwasowego pożywki lub układu reakcyjnego | Selektywnie obniża pulę L-asparaginy | Nie zastępuje kontroli całego metabolizmu azotu |
| Badania metabolizmu komórkowego | Modelowanie deprywacji L-asparaginy | Tworzy warunki niskiej dostępności tego aminokwasu | Nie jest równoważna pełnemu usunięciu wszystkich źródeł azotu |
| Rozwój enzymów i biokatalizy | Poszukiwanie wariantów o innym profilu stabilności lub selektywności | Dostępne są liczne naturalne i inżynieryjne warianty L-asparaginaz | Dane z jednego wariantu nie powinny być automatycznie przenoszone na inny |
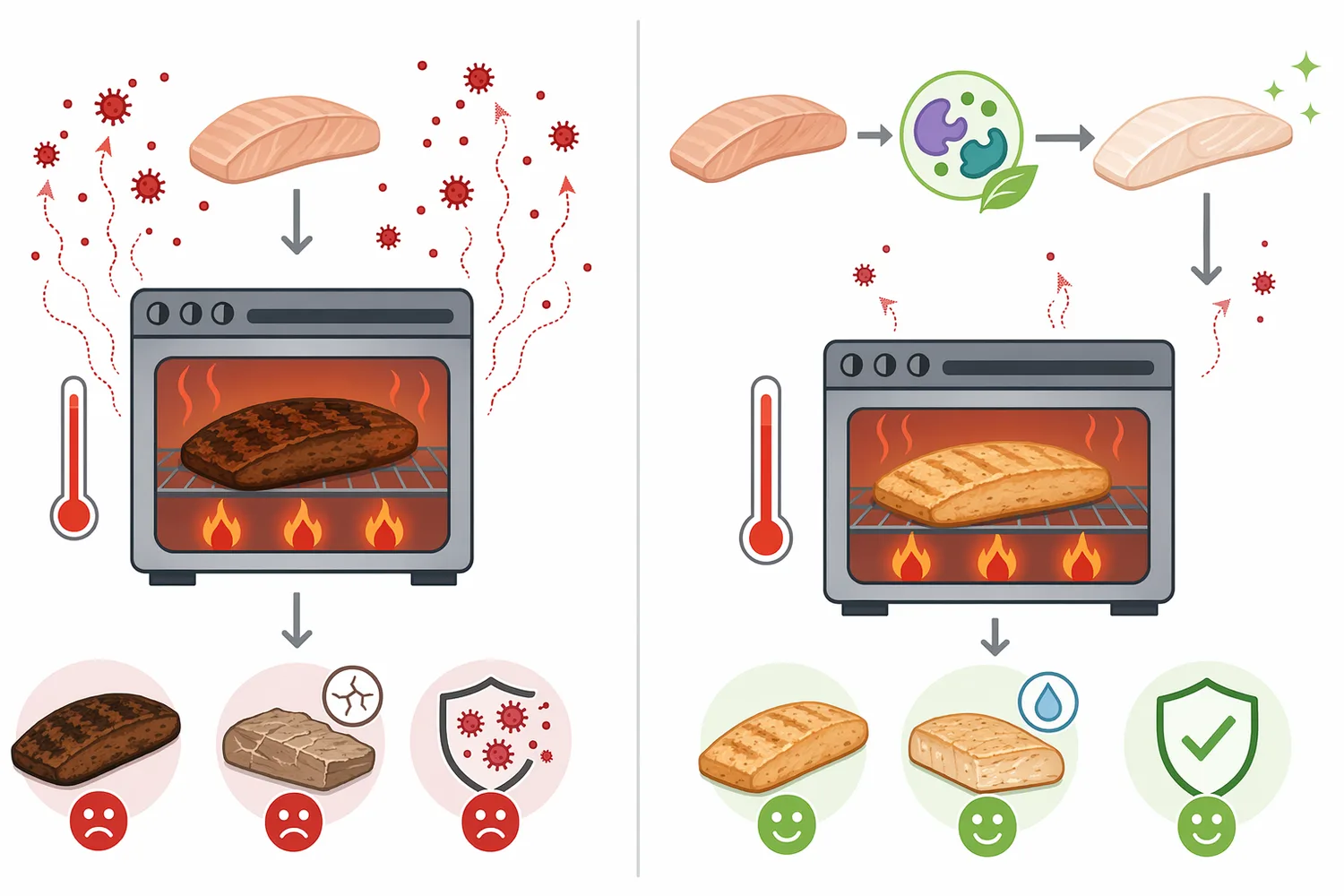
W technologii żywności L-asparaginase jest kojarzona przede wszystkim z ograniczaniem akrylamidu w produktach poddawanych wysokiej temperaturze. Mechanizm jest racjonalny i bezpośredni: enzym nie „neutralizuje” akrylamidu po jego powstaniu, lecz działa wcześniej, zmniejszając pulę L-asparaginy dostępnej do reakcji z cukrami redukującymi [2].
Z punktu widzenia procesu kluczowe jest wprowadzenie enzymu na etapie, na którym substrat jest jeszcze dostępny, a środowisko nie dezaktywuje białka zbyt szybko. Dlatego asparaginase rozważa się w fazach uwodnionych, mieszanych lub takich, w których można zapewnić kontakt enzymu z wolną L-asparaginą przed pieczeniem, smażeniem, prażeniem lub innym etapem wysokotemperaturowym [2].
Nie oznacza to, że efekt będzie identyczny w każdej recepturze. Zawartość L-asparaginy różni się między surowcami, a aktywność enzymatyczna zależy od pH, temperatury, czasu, dyfuzji w matrycy i obecności składników ograniczających dostęp do substratu. Dlatego w dokumentacji technicznej należy mówić o mechanizmie redukcji prekursora, a nie o uniwersalnej, stałej redukcji końcowego parametru w każdej aplikacji [1].
W biotechnologii asparaginase może być używana jako narzędzie do selektywnego obniżania poziomu L-asparaginy w pożywkach, hydrolizatach lub układach modelowych. W odróżnieniu od metod nieselektywnych, które zmieniają całkowitą pulę aminokwasów, L-asparaginase wpływa przede wszystkim na jeden związek i przekształca go w L-asparaginian oraz amoniak [1].
Taki mechanizm jest przydatny w badaniach metabolizmu azotu, zależności wzrostu od aminokwasów oraz odpowiedzi komórek na ograniczenie dostępności L-asparaginy. W literaturze L-asparaginase jest szeroko omawiana jako enzym wywołujący deprywację asparaginy, co pozwala badać komórki lub organizmy zależne od zewnętrznej podaży tego aminokwasu [4].
Jednocześnie trzeba pamiętać, że L-asparaginase nie usuwa całego azotu organicznego i nie blokuje wszystkich dróg biosyntezy aminokwasów. Jeżeli układ biologiczny może syntetyzować L-asparaginę z innych prekursorów, sama degradacja wolnej puli substratu może wywołać efekt przejściowy lub zależny od dynamiki metabolizmu [1].
L-asparaginase jest jednym z najlepiej znanych enzymów przeciwnowotworowych w historii biochemii klinicznej, szczególnie w kontekście ostrej białaczki limfoblastycznej. Mechanizm terapeutyczny opiera się na deprywacji L-asparaginy: komórki wrażliwe na jej niedobór tracą dostęp do aminokwasu potrzebnego do syntezy białek i utrzymania wzrostu [1].
Ta wiedza jest istotna dla zrozumienia enzymu, ale nie powinna być mylona z przeznaczeniem produktu dostarczanego do zastosowań B2B. Informacje o aktywności przeciwnowotworowej L-asparaginaz opisanych w badaniach naukowych dotyczą określonych białek, formulacji, systemów jakości i kontekstów regulowanych; nie stanowią automatycznej deklaracji zastosowania dowolnego produktu asparaginase w terapii, diagnostyce lub podaniu ludziom [4].
W pracach badawczych porównywano również L-asparaginazy z różnych źródeł pod kątem właściwości biologicznych i biochemicznych. Przykładowo enzymy z izolowanych szczepów Escherichia coli były oceniane w modelach komórkowych, co pokazuje znaczenie pochodzenia enzymu, jego profilu aktywności i właściwości ubocznych dla interpretacji wyników [4].
L-asparaginase nie jest pojedynczą, identyczną cząsteczką występującą we wszystkich produktach i organizmach. Enzymy o tej funkcji opisano u bakterii, cyjanobakterii, drożdży i innych mikroorganizmów, a ich właściwości mogą się różnić pod względem stabilności, preferencji pH, temperatury działania, oligomeryzacji i aktywności ubocznej wobec L-glutaminy [2].
Badania nad L-asparaginase z Synechococcus elongatus pokazują, że także cyjanobakterie mogą być źródłem wariantów o interesujących właściwościach biochemicznych i aktywności biologicznej. Takie prace są ważne dla rozwoju biokatalizy, ponieważ poszerzają pulę enzymów poza klasyczne źródła bakteryjne wykorzystywane historycznie [5].

Opisano również rekombinowaną L-asparaginase pochodzącą z morskiego szczepu Pseudomonas aeruginosa, wraz z analizą strukturalną i biochemiczną. Dla zastosowań procesowych znaczenie ma tu nie sam fakt „morskiego” pochodzenia, lecz to, że enzymy z różnych nisz ekologicznych mogą wykazywać odmienne profile stabilności i aktywności [6].
Źródłem L-asparaginase mogą być także drożdże izolowane ze środowiska glebowego. Badania nad rodzimymi izolatami z gleby wskazują, że poszukiwanie nowych enzymów nie ogranicza się do standardowych mikroorganizmów laboratoryjnych, a różnorodność biologiczna pozostaje ważnym zasobem dla biotechnologii enzymów [7].
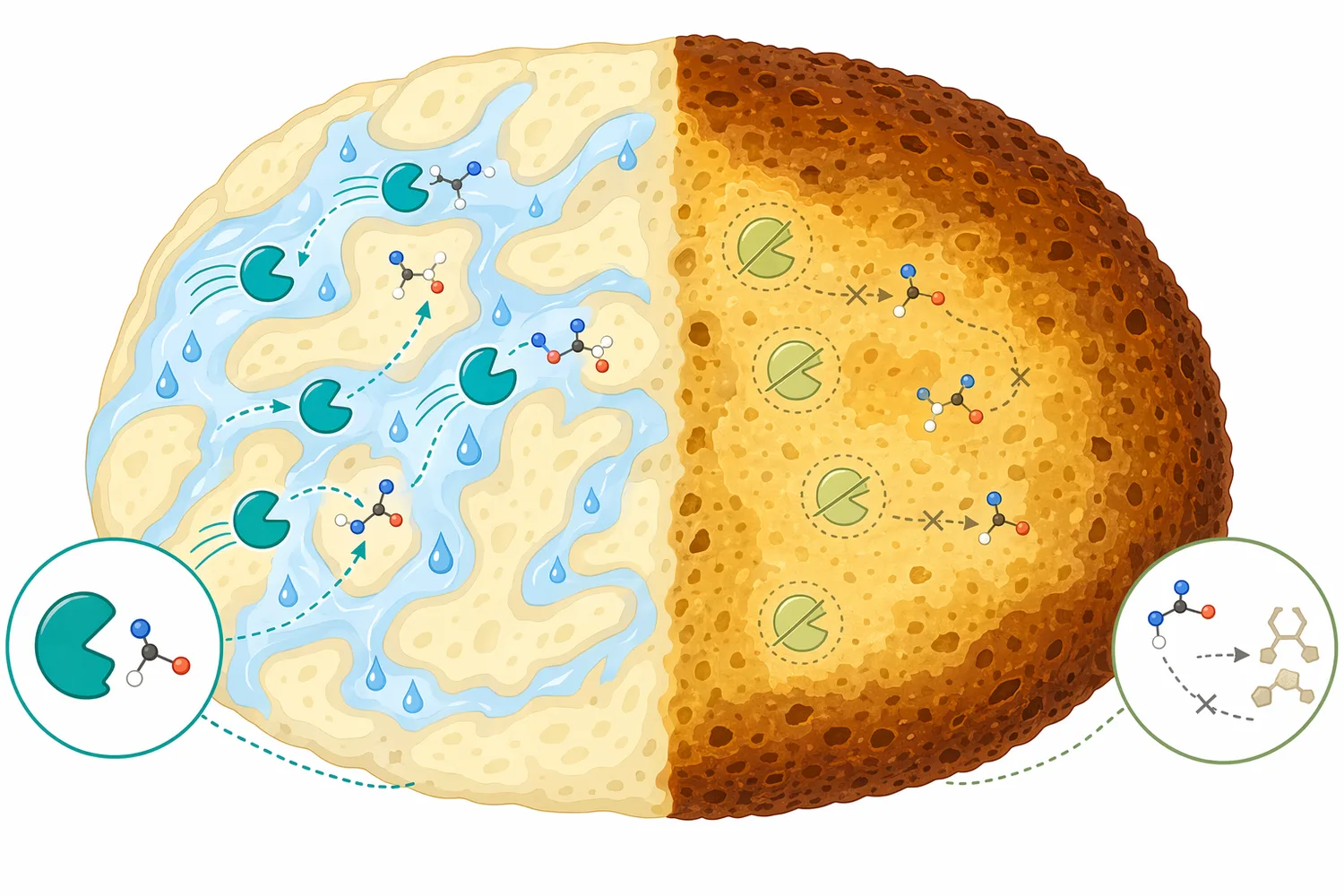
Najważniejszym warunkiem skuteczności asparaginase jest dostępność L-asparaginy. Enzym działa na substrat, z którym ma kontakt; jeśli aminokwas jest fizycznie uwięziony w strukturze surowca, słabo rozpuszczony, związany w niedostępnej frakcji albo obecny dopiero po późniejszym etapie hydrolizy białek, reakcja może być ograniczona [1].
Drugim czynnikiem jest woda, ponieważ reakcja jest hydrolizą. W matrycach o niskiej aktywności wody reakcje enzymatyczne zwykle przebiegają wolniej, a białko może mieć ograniczoną ruchliwość konformacyjną potrzebną do katalizy. Dlatego asparaginase najczęściej rozważa się tam, gdzie istnieje faza wodna lub przynajmniej etap wystarczającego uwodnienia [2].
Trzecim elementem jest pH. W literaturze dla różnych L-asparaginaz mikrobiologicznych opisywano optima w zakresie od lekko kwaśnego do zasadowego, często w pobliżu warunków neutralnych lub umiarkowanie alkalicznych, ale nie istnieje jedna wartość właściwa dla wszystkich wariantów enzymu. Różnice wynikają z sekwencji białka, architektury centrum aktywnego i pochodzenia organizmu [2].
Czwartym elementem jest temperatura. Wzrost temperatury może przyspieszać reakcję do momentu, w którym zaczyna dominować utrata struktury białkowej; dlatego „cieplej” nie znaczy automatycznie „lepiej”. W procesach z późniejszą obróbką cieplną enzym powinien zadziałać przed etapem, który może go trwale zdezaktywować [1].
Piątym czynnikiem jest skład matrycy. Sole, cukry, białka, związki fenolowe, środki redukujące, metale i produkty uboczne mogą wpływać na stabilność oraz dostępność substratu. Przykładowo badania nad izoformami L-asparaginase z Rhizobium etli wskazują, że jony metali — w tym cynk — mogą istotnie zmieniać właściwości biochemiczne niektórych enzymów, co pokazuje, jak silnie kontekst chemiczny może modulować działanie [8].
W zastosowaniach technicznych często zakłada się, że asparaginase jest selektywna wobec L-asparaginy, ale w praktyce selektywność należy rozumieć jako cechę konkretnego enzymu, a nie całej nazwy handlowej. Niektóre L-asparaginazy wykazują aktywność wobec L-glutaminy, a inne są projektowane lub selekcjonowane w kierunku mniejszej aktywności ubocznej [1].
Dlaczego to ma znaczenie? L-glutamina jest również ważnym aminokwasem amidowym i w wielu układach biologicznych pełni funkcje odżywcze, buforujące oraz metaboliczne. Jeżeli proces wymaga wyłącznie redukcji L-asparaginy, nadmierna aktywność glutaminazowa może nie być pożądana, ponieważ zmienia szerszy profil aminokwasowy matrycy [1].
Współczesne badania nad L-asparaginase obejmują zatem nie tylko poszukiwanie wysokiej aktywności, ale także modyfikację stabilności, selektywności i właściwości ubocznych. Prace wykorzystujące rekonstrukcję sekwencji ancestralnych pokazują, że inżynieria białek może prowadzić do wariantów o ulepszonych cechach w porównaniu z enzymami wyjściowymi [9].
Podobnie badania nad podstawieniami aminokwasowymi inspirowanymi naturą w EcAIII pokazują, że nawet pojedyncze zmiany sekwencji mogą wpływać na właściwości strukturalne i biochemiczne enzymu. To ważna lekcja praktyczna: nie należy przenosić parametrów jednego wariantu L-asparaginase na inny bez weryfikacji w danym układzie [3].
Asparaginase jest dobrym kandydatem do prac rozwojowych, gdy problem da się sformułować wprost: „potrzebujemy zmniejszyć ilość L-asparaginy”. Jeśli celem jest obniżenie dostępności tego aminokwasu w matrycy, pożywce lub półprodukcie, mechanizm enzymu odpowiada zadaniu bezpośrednio [1].
Nie jest natomiast właściwym narzędziem, gdy źródło problemu leży gdzie indziej: w ogólnej zawartości białka, niepożądanej proteolizie, utlenianiu lipidów, zanieczyszczeniu mikrobiologicznym, nieodpowiedniej lepkości lub obecności innych aminokwasów. W takich przypadkach zastosowanie L-asparaginase może nie przynieść mierzalnej korzyści, ponieważ enzym nie oddziałuje na pierwotną przyczynę [2].
W badaniach metabolizmu enzym pozwala zaprojektować warunki ograniczonej dostępności L-asparaginy bez całkowitego usuwania innych składników pożywki. To szczególnie przydatne, gdy zespół chce rozdzielić efekt deprywacji jednego aminokwasu od ogólnego stresu głodowego lub od zmian wynikających z całkowitej przebudowy medium [4].
W procesach żywnościowych i technologicznych asparaginase najlepiej sprawdza się jako element strategii redukcji prekursora, a nie jako późny środek naprawczy. Jeżeli niepożądany produkt już powstał, enzym hydrolizujący L-asparaginę nie cofnie wcześniejszej reakcji; jego rola polega na zmniejszeniu ryzyka powstania produktu przez ograniczenie substratu przed etapem krytycznym [2].
Najważniejsze ograniczenie jest proste: L-asparaginase działa tylko wtedy, gdy obecna jest L-asparagina w formie dostępnej dla enzymu. Jeżeli substrat jest nieobecny albo jego ilość nie wpływa na problem technologiczny, zastosowanie enzymu nie ma logicznego uzasadnienia [1].
Drugie ograniczenie dotyczy stabilności białka. Enzymy są katalizatorami biologicznymi, a ich aktywność zależy od struktury trzeciorzędowej i — w wielu przypadkach — czwartorzędowej. Skrajne pH, długie ogrzewanie, niezgodne rozpuszczalniki, wysokie stężenia niektórych soli lub składniki denaturujące mogą ograniczać czas efektywnego działania [1].
Trzecie ograniczenie dotyczy przenoszenia wniosków między branżami. Fakt, że L-asparaginase jest intensywnie badana w onkologii, nie oznacza, że każdy produkt enzymatyczny jest odpowiedni do zastosowań terapeutycznych. Badania biomedyczne dotyczą określonych enzymów, oczyszczania, formulacji, dawek, reżimów jakości i oceny bezpieczeństwa w ramach regulowanych systemów [4].
Czwarte ograniczenie dotyczy matrycy. W literaturze opisano liczne L-asparaginazy o różnych właściwościach, w tym warianty pochodzące z bakterii, cyjanobakterii, drożdży i organizmów patogennych. Na przykład sekrecyjna L-asparaginase Rv1538c z Mycobacterium tuberculosis była badana w kontekście przeżycia w fagosomie, co podkreśla, że funkcja enzymu może być osadzona w bardzo różnym kontekście biologicznym [10].
Dane o L-asparaginase warto interpretować na trzech poziomach. Pierwszy poziom to reakcja chemiczna: hydroliza L-asparaginy do L-asparaginianu i amoniaku. Ten mechanizm jest centralny, dobrze opisany i wspólny dla zastosowań enzymu [1].
Drugi poziom to właściwości konkretnego białka: struktura, oligomeryzacja, profil substratowy, stabilność i odpowiedź na warunki środowiska. Tu różnice między enzymami mogą być duże, dlatego publikacja dotycząca jednego wariantu nie opisuje automatycznie wszystkich produktów określanych jako asparaginase [3].
Trzeci poziom to wynik aplikacyjny w matrycy. Redukcja L-asparaginy może przełożyć się na pożądany efekt technologiczny, ale skala tego efektu zależy od procesu, receptury i etapu dodania enzymu. Właśnie dlatego w zastosowaniach B2B najbezpieczniej traktować asparaginase jako narzędzie mechanistyczne: bardzo użyteczne, gdy problem dotyczy L-asparaginy, ale wymagające dopasowania do realnych warunków produkcyjnych [2].
Enzymes.bio jest dostawcą enzymów B2B, a nie producentem ani laboratorium badawczym. Asparaginase jest oferowana online w jednostkach 1 kg, a zamówienie jest realizowane po płatności online. Dokumenty CoA i SDS są dostarczane wraz z zamówieniem.
W kontekście dokumentacji technicznej najważniejsze jest odpowiedzialne przypisanie roli produktu: asparaginase służy do zastosowań, w których uzasadnione jest kontrolowane obniżanie poziomu L-asparaginy. Wszelkie zastosowania regulowane — w tym żywnościowe, farmaceutyczne, kosmetyczne, diagnostyczne lub badawcze objęte szczególnymi wymaganiami — powinny być oceniane przez użytkownika końcowego zgodnie z lokalnymi przepisami i przeznaczeniem danego procesu.
Asparaginase, czyli L-asparaginase, jest precyzyjnym enzymem do deamidacji L-asparaginy. Jej podstawowa reakcja przekształca L-asparaginę w L-asparaginian i amoniak, co pozwala obniżyć dostępność tego aminokwasu w układach technologicznych, biologicznych i badawczych [1].
Najbardziej praktyczne zastosowania wynikają z tej samej zasady: jeśli L-asparagina jest niepożądanym prekursorem albo czynnikiem wpływającym na wzrost, metabolizm lub jakość produktu, enzymatyczna konwersja może być logicznym rozwiązaniem. W technologii żywności oznacza to między innymi redukcję prekursora akrylamidu przed etapami wysokotemperaturowymi; w biotechnologii i R&D — kontrolę dostępności konkretnego aminokwasu [2].
Jednocześnie skuteczność asparaginase nie jest cechą oderwaną od warunków. Decydują dostępność substratu, woda, pH, temperatura, czas kontaktu, skład matrycy i właściwości konkretnego wariantu enzymu. Dlatego najlepiej rozumieć L-asparaginase jako ukierunkowane narzędzie do problemów związanych z L-asparaginą — nie jako uniwersalny enzym procesowy do każdej matrycy.
Sprzedawany w jednostkach 1 kg, dostępny z magazynu i gotowy do wysyłki. Zamów bezpośrednio w naszym sklepie — zapłać online, a my przetworzymy Twoje zamówienie. Do każdego zamówienia dołączamy Certyfikat Analizy i Kartę Charakterystyki.
Kup Asparaginase →Ponumerowano według kolejności pierwszego cytowania. Źródła open access, każde zweryfikowane jako dostępne w momencie publikacji; numery cytowań w tekście prowadzą tutaj.