Beta-amylase is a maltose-forming starch enzyme: it works from the non-reducing ends of accessible starch chains and releases maltose units rather than randomly cutting the chain throughout. That makes it especially useful when the process target is a maltose-rich sugar profile, such as in brewing, distilling, cereal fermentation, and high-maltose syrup production. Enzymes.bio supplies Beta-Amylase directly online by the 1 kg unit; buyers place and pay for the order online, and the order is then processed and shipped with a Certificate of Analysis and Safety Data Sheet.
Beta-amylase, also written as beta amylase or β-amylase, is an amylolytic enzyme that converts starch-derived chains into maltose-rich hydrolysates. Its defining feature is exo-action: instead of attacking a starch molecule at random internal positions, it begins at a non-reducing chain end and removes maltose, a two-glucose sugar, step by step. Early action-pattern studies on amylose established this multichain, end-wise mode of attack, which remains central to how the beta amylase enzyme is understood in starch conversion today [1].
This action is highly specific to the structure of starch. Starch contains glucose units linked mainly by α-1,4 glucosidic bonds in linear regions, with α-1,6 branch points in amylopectin. Beta-amylase hydrolyzes the α-1,4 bonds accessible from non-reducing ends, so the visible process effect is not simply “starch disappears”; rather, the carbohydrate profile shifts toward maltose while larger residual dextrins remain when the enzyme reaches a branch point or an inaccessible segment [2].
The name can be confusing because the “beta” in beta-amylase does not mean it hydrolyzes beta-linked starch. Starch is still an α-glucan substrate; the enzyme is called beta-amylase because the maltose product is released in the beta anomeric form. Mechanistic work on beta-amylase reactions with maltosyl fluoride substrates helped clarify that beta-amylase chemistry is tied to the stereochemical outcome of maltose release, not to a beta-linked starch substrate [3].
In practical terms, beta-amylase is valuable when a processor wants maltose rather than a broad mixture of dextrins, glucose, and short oligosaccharides. Maltose is readily fermentable by many brewing and distilling yeasts, and it is also the defining sugar in high-maltose syrups. For customers searching “what is beta-amylase,” the most useful answer is therefore process-oriented: it is the enzyme used to turn accessible starch chain ends into maltose-rich sugar streams.
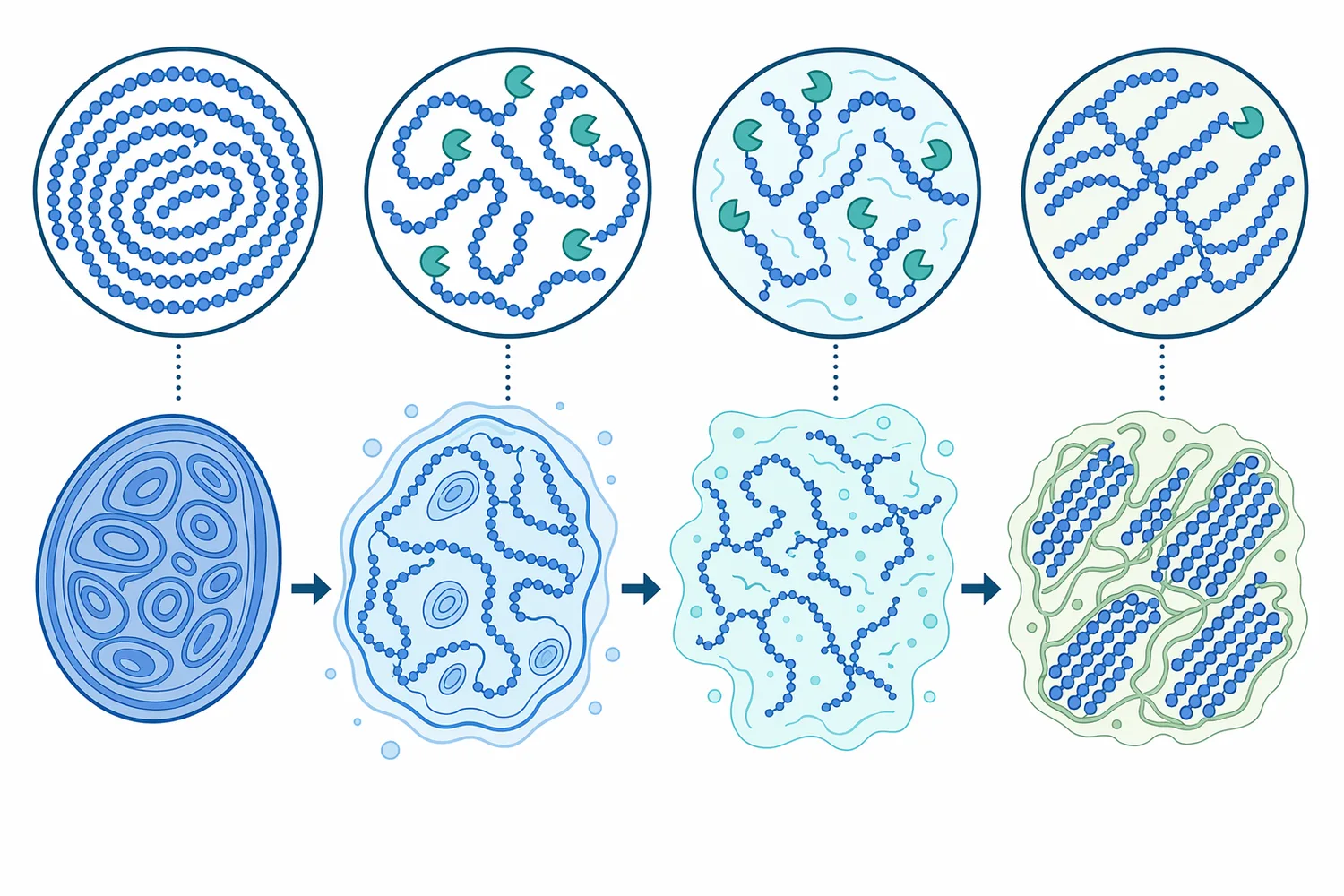
A starch chain can be visualized as a long row of glucose units. Beta-amylase binds near a non-reducing end, positions an α-1,4 bond in its catalytic site, adds water across that bond, and releases maltose. After one maltose unit is removed, a new non-reducing end is exposed, allowing the enzyme to continue along the same chain or move to another chain; classic studies described this as multichain action rather than single complete digestion of one molecule before moving on [4].
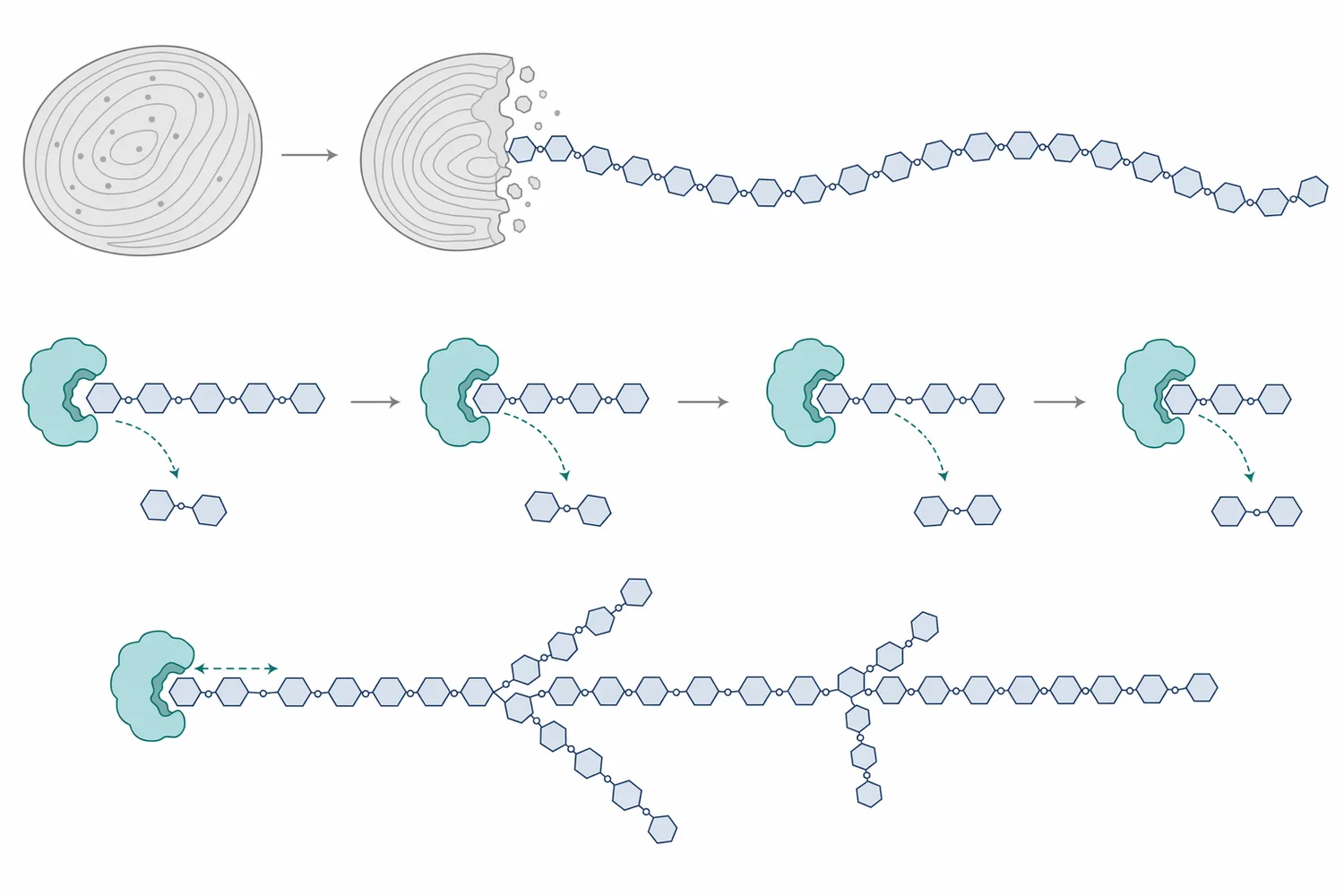
Later kinetic and structural analysis added a more detailed picture: beta-amylase can perform multiple attacks during one productive encounter with a starch chain. In other words, after releasing one maltose, the enzyme-substrate complex may reposition so that another bond can be cleaved without complete diffusion away from the substrate. This “sliding” behavior helps explain why beta-amylase can efficiently generate maltose from accessible amylose-like regions while still being limited by branch points and steric constraints [5].
The branch-point limitation is important in real starch systems. Amylose is mostly linear, so beta-amylase can remove maltose repeatedly along much of the chain. Amylopectin is branched, so beta-amylase proceeds along an outer chain until it approaches an α-1,6 branch point; the enzyme does not simply cut through that branch and continue. The remaining branched material is commonly described as beta-limit dextrin, because it represents the practical limit of beta-amylase action on branched starch under those conditions [2].
This mechanism explains why beta-amylase is often used after starch has already been cooked, gelatinized, milled, malted, or partially liquefied. Native starch granules protect much of the glucan structure inside crystalline and semi-crystalline regions, so the number of accessible chain ends is limited. When heat, water, milling, germination, or alpha-amylase liquefaction opens the granule and reduces viscosity, beta-amylase has more exposed chain ends to work on, and maltose formation becomes more efficient.
The common comparison “alpha vs beta amylase” matters because the two enzymes solve different starch-processing problems. Alpha-amylase is endo-acting: it cuts internal α-1,4 bonds within starch chains, rapidly reducing viscosity and producing a mixture of dextrins and shorter oligosaccharides. Beta-amylase is exo-acting: it works from non-reducing ends and releases maltose in a more ordered, maltogenic pattern. Studies on alpha-amylase action patterns and active-site catalysis support this distinction between internal chain cleavage and the chain-end maltose release associated with beta-amylase [6].
| Enzyme type | Main cutting pattern | Typical process effect | Main practical role |
|---|---|---|---|
| Alpha-amylase | Internal α-1,4 cleavage | Rapid viscosity reduction; mixed dextrins and oligosaccharides | Liquefaction, thinning, preparation of starch for further saccharification |
| Beta-amylase | Non-reducing-end α-1,4 cleavage | Maltose-rich hydrolysate; beta-limit dextrins remain near branch points | Maltose formation for brewing, fermentation, and high-maltose syrups |
| Glucoamylase / gamma-amylase terminology | Exo-release of glucose from chain ends | Higher glucose formation; different sugar profile from beta-amylase | Glucose-rich saccharification rather than maltose-focused conversion |
For readers comparing alpha amylase vs beta amylase, the core difference is therefore not “stronger versus weaker.” It is “liquefying versus maltose-forming.” Alpha-amylase opens the starch structure by creating more shorter chains, while beta-amylase turns accessible non-reducing ends into maltose. In many processes, alpha amylase and beta amylase are complementary rather than interchangeable, especially when a process first needs viscosity reduction and then controlled maltose production.
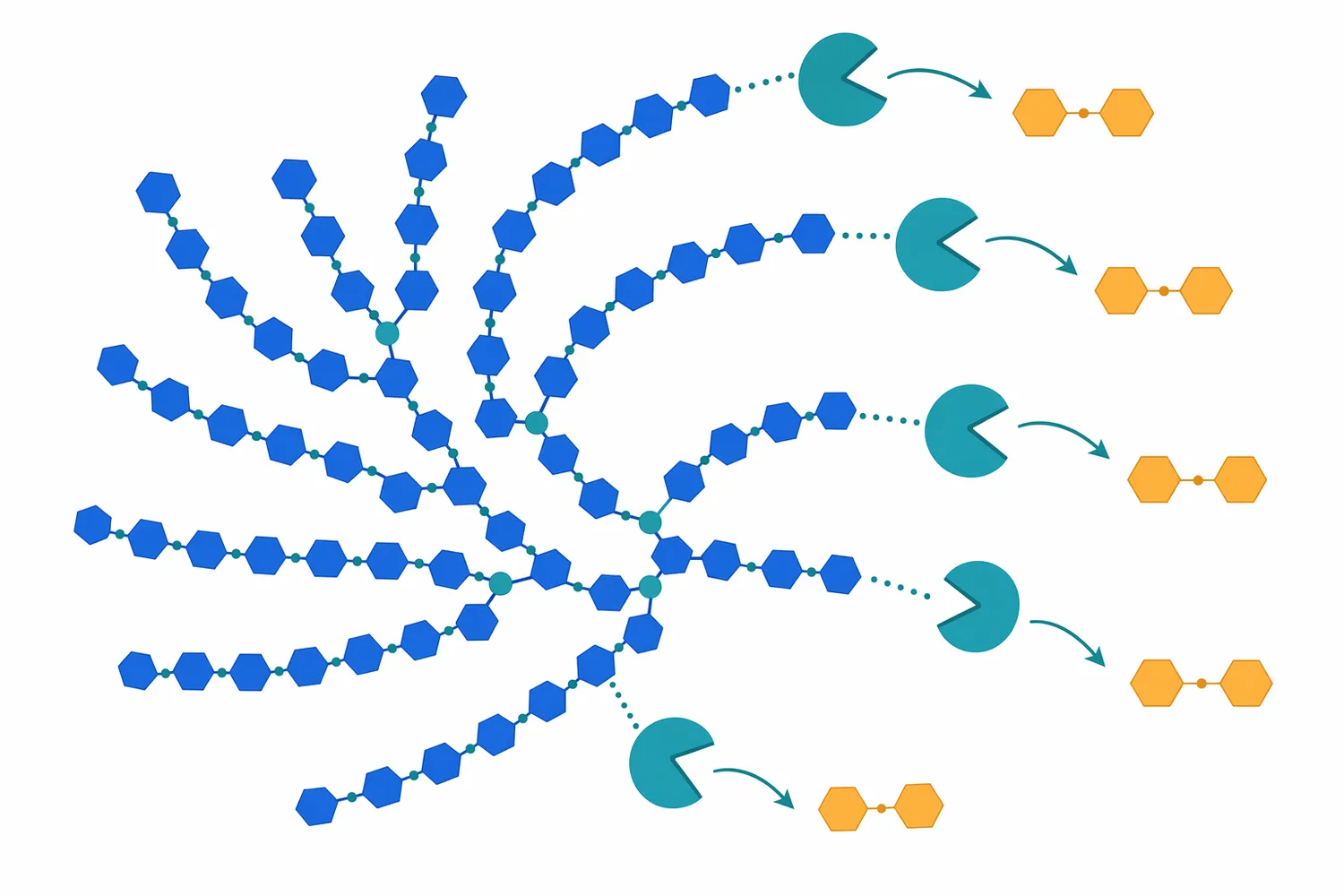
The phrase “alpha beta and gamma amylase” is sometimes used in broad searches, but the practical question is the desired sugar profile. Alpha-amylase tends to broaden the dextrin pool, beta-amylase enriches maltose, and glucose-forming exoamylases shift the product toward glucose. For a buyer interested in beta amylase enzyme for sale, the relevant application fit is therefore the need for maltose-rich conversion, not simply the need for any starch-degrading enzyme.
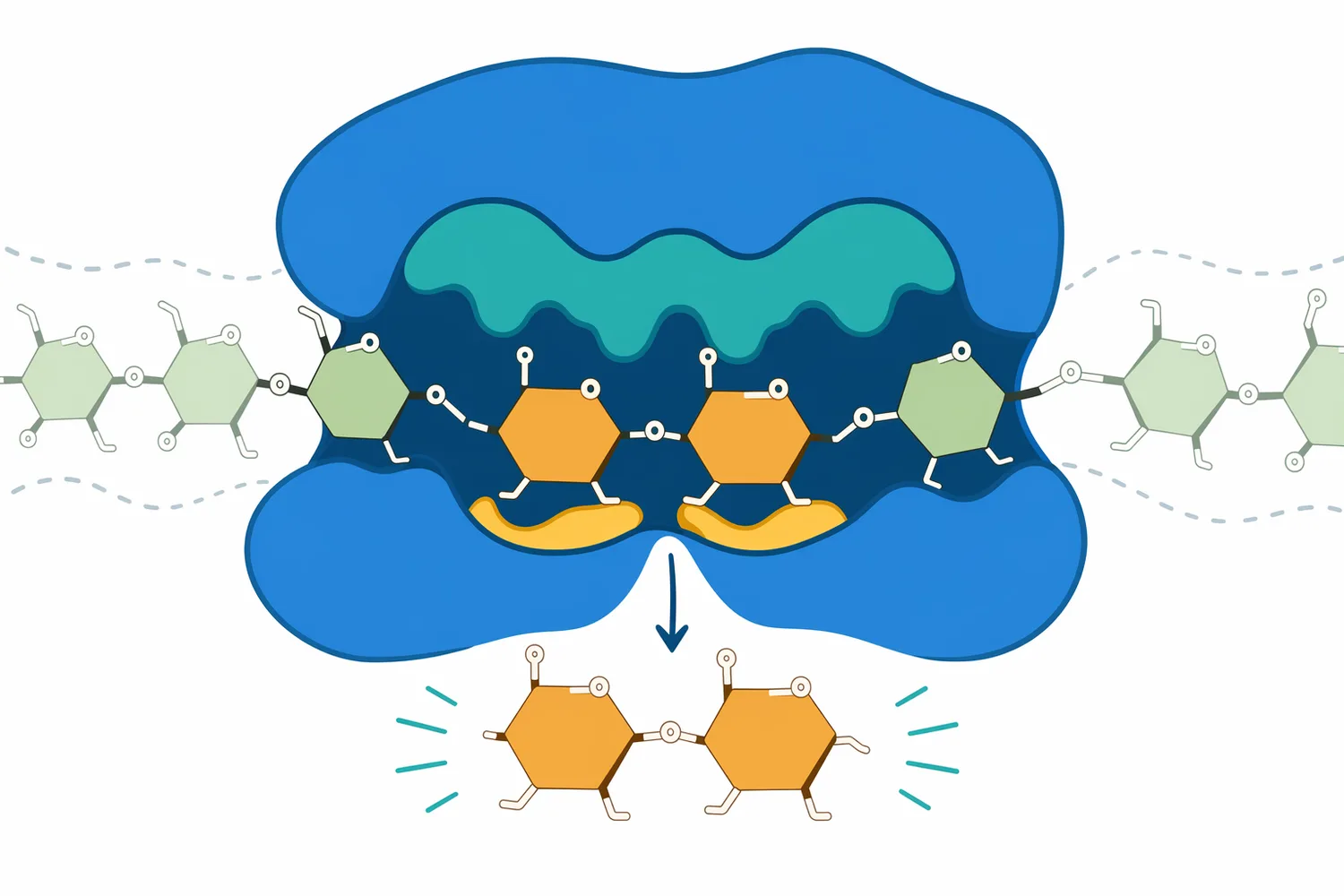
Beta-amylase structure is built around a catalytic site that accommodates a starch chain end in a defined orientation. The enzyme must recognize the non-reducing end, align the correct α-1,4 bond, and position catalytic residues so that hydrolysis releases maltose rather than a random-length fragment. Functional analysis of soybean beta-amylase identified the importance of specific residues, including Glu380 and Leu383, in the proposed catalytic mechanism and substrate handling [7].
That active-site geometry is what makes beta-amylase selective. The enzyme is not simply a chemical acid that breaks glycosidic bonds wherever it contacts starch; it is a folded protein that uses binding subsites to hold glucose residues in a productive arrangement. If the chain is too close to an α-1,6 branch point, too short, poorly accessible, or trapped inside ungelatinized granule structure, the enzyme cannot maintain the same productive progression.
The structural view also explains the importance of multiple attack. When beta-amylase remains associated with a chain and slides to the next cleavage position, it can release successive maltose units efficiently. But this process still depends on the substrate remaining aligned and accessible; if branching, retrogradation, protein-starch interactions, or processing conditions restrict access, maltose release slows even if starch is still present [5].
This is why beta-amylase performance is best understood as a combination of enzyme chemistry and substrate architecture. The enzyme’s catalytic identity determines what it can make—mainly maltose from α-1,4 linked chain ends. The raw material and process history determine how much of the starch is actually available for that action.
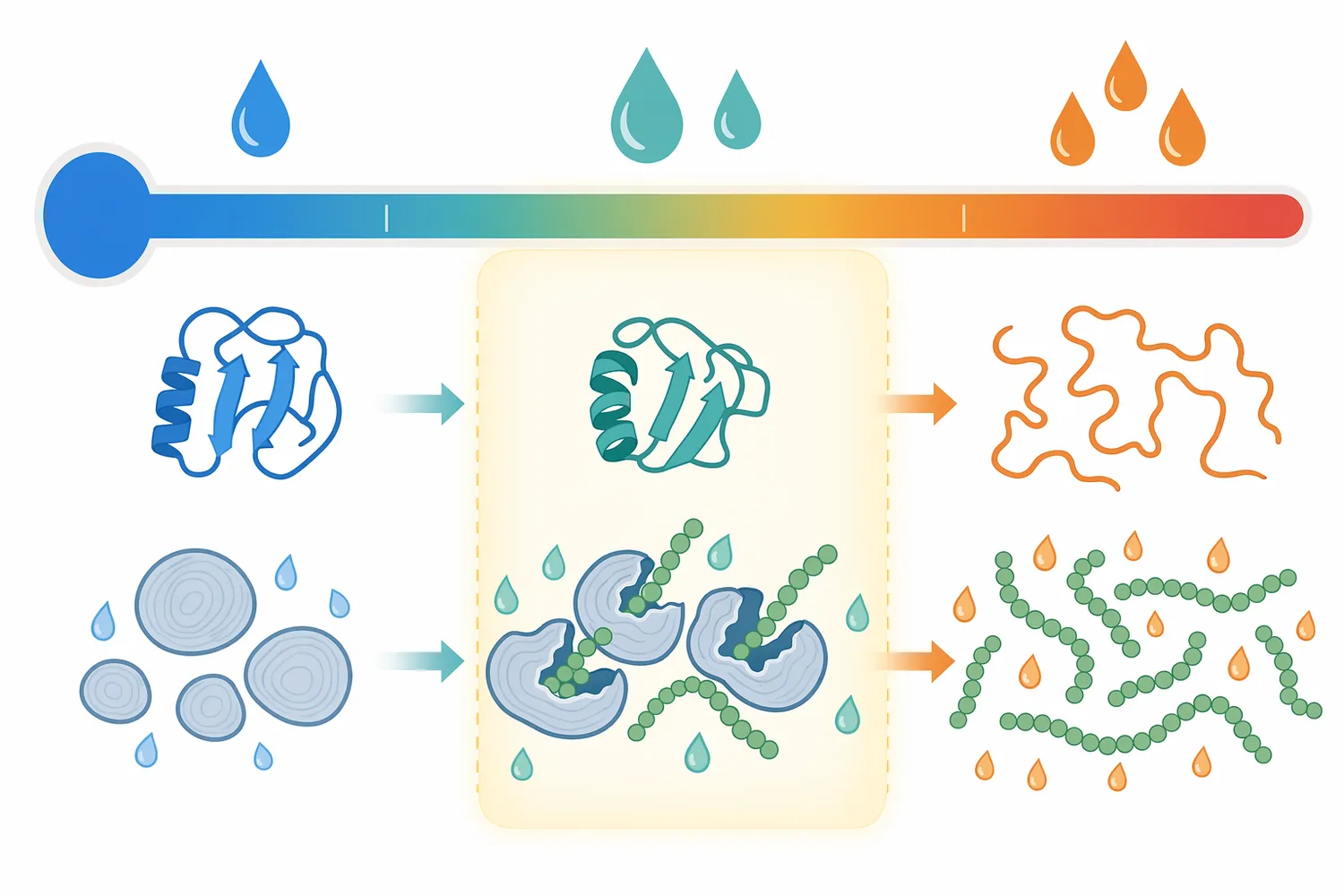
The beta amylase temperature range discussed in processing literature is usually moderate compared with high-temperature liquefaction steps. In brewing and cereal processing, beta-amylase is commonly associated with mash or saccharification conditions rather than the hottest starch-cooking stages. The reason is structural: enzymes are proteins, and excessive heat can unfold the active conformation needed to bind chain ends and catalyze maltose release.
A useful way to frame temperature is by function. If the starch is not hydrated and opened, beta-amylase may have few accessible chain ends even if the enzyme itself remains active. If the temperature is high enough to damage the enzyme, accessibility may improve but catalytic capacity falls. The process window is therefore the overlap between starch accessibility and enzyme stability, which is why beta-amylase is often used after upstream steps have made starch more accessible.
Plant beta-amylases have been widely studied because cereals and germinating seeds naturally mobilize starch reserves. Immunochemical work on germinating rice seeds showed that amylase systems can include multiple forms during starch breakdown, reinforcing that biological starch mobilization is not a single-enzyme event but a coordinated enzymatic process [8]. For industrial users, the same principle applies: beta-amylase contributes a specific maltose-forming function within a broader starch-conversion workflow.
Source also matters. Classic mechanistic studies include plant enzymes such as soybean beta-amylase, while other enzyme preparations in biotechnology may be microbial. The search phrase “beta amylase producing bacteria” reflects interest in microbial enzyme origins, but application performance still comes back to the same practical questions: whether the enzyme can access the substrate, remain folded under the process conditions, and release maltose at the desired stage of conversion. Microbial enzyme preparations are broadly important across biotechnological industries, including starch-processing contexts [9].
Beta amylase beer applications are among the most familiar uses of the enzyme. During mashing, malted grain starch is hydrated and enzymatically converted into soluble sugars. Beta-amylase helps generate maltose, which brewing yeast can ferment, while alpha-amylase helps open starch chains and produce dextrins that influence body and residual extract. The balance of alpha and beta amylase brewing activity therefore affects fermentability, mouthfeel, and the final carbohydrate profile of the wort.
In malt, beta-amylase is part of the natural enzyme system developed during germination. The enzyme acts on starch fragments and exposed chain ends created during mashing, steadily enriching the wort in maltose. If beta-amylase activity is limited by raw material, heat exposure, or insufficient substrate accessibility, the wort may retain more higher oligosaccharides and less maltose than expected.
The practical distinction between alpha and beta amylase is especially important in brewing because mash programs are partly designed around enzyme behavior. A hotter, more liquefying-oriented mash favors rapid starch thinning and dextrin formation; a more beta-amylase-friendly saccharification rest supports maltose production. This does not mean beta-amylase works alone—rather, it contributes the maltose-forming part of a multi-enzyme starch conversion system.
For distilling and cereal-based fermentation, the same chemistry applies. Yeast does not efficiently consume intact starch, so the substrate must be converted into fermentable sugars before or during fermentation. Beta-amylase contributes maltose formation, while the final fermentable profile depends on the extent of starch opening, the presence of complementary enzymes, and how long the substrate remains in conditions that preserve beta-amylase function.
High-maltose syrup production is a natural fit for beta-amylase because maltose is the enzyme’s principal product. After starch is cooked and opened, beta-amylase can remove maltose units from exposed non-reducing ends, shifting the syrup composition toward a maltose-rich profile. This is different from a glucose-syrup strategy, where the process is designed to push conversion toward glucose rather than maltose.
The important limitation is amylopectin branching. Corn, wheat, potato, tapioca, and other starches contain different amylose-to-amylopectin ratios and different granule structures, but any amylopectin-rich starch presents branch points that beta-amylase cannot simply pass. Once the enzyme approaches an α-1,6 branch, maltose release from that chain segment stops, leaving beta-limit dextrin. The observed syrup composition is therefore a direct reflection of enzyme specificity plus starch structure [2].
In many starch-conversion systems, beta-amylase is paired conceptually with enzymes that perform other tasks. Alpha-amylase can reduce viscosity and create more chain ends; debranching enzymes can expose additional linear segments; glucose-forming enzymes can change the final sweetness and fermentability profile. Beta-amylase remains the maltose-forming component in that system, and its value is the predictability of its product pattern.
This is why the phrase “beta alpha amylase” sometimes appears in searches, even though the more precise wording is alpha and beta amylase. The two enzyme classes are not duplicates. Alpha-amylase improves access by internal cleavage, while beta-amylase converts accessible ends into maltose. When the desired result is a maltose-rich syrup, beta-amylase provides the product direction that alpha-amylase alone does not.
In cereal doughs, porridges, mashes, and fermented grain systems, beta-amylase contributes to sugar release from starch fragments. The effect is especially relevant when maltose supports yeast activity or when the product profile benefits from a mild, maltose-forward sweetness. Because cereal matrices contain starch, proteins, fibers, lipids, and minerals, the actual conversion pattern depends on how processing exposes starch and how competing physical effects influence enzyme access.
Germinating seeds provide a useful natural model. During germination, stored starch must be mobilized into smaller sugars that support plant growth, and amylase systems become active as part of that mobilization. Research on rice seed amylases showed multiple forms during germination, highlighting that cereal starch breakdown involves coordinated enzymes rather than one universal amylase [8].
Mustard cotyledon beta-amylase research likewise illustrates the plant-enzyme context of beta-amylase biology. Studies of beta-amylase from plant cotyledons are relevant because they connect the enzyme’s natural role—mobilizing stored carbohydrate—to its industrial role—converting accessible starch into maltose-rich hydrolysate [10].
For food fermentation, the practical outcome is not only sugar quantity but sugar type. Maltose can support yeast metabolism, influence fermentation kinetics, and contribute to the balance between fermentable sugars and residual carbohydrates. Beta-amylase is therefore most useful where maltose formation is part of the desired sensory, fermentation, or syrup profile.
A common processing mistake is to think of starch as a uniform soluble substrate. In reality, starch exists in granules, swollen granules, gelatinized paste, liquefied dextrin solution, or retrograded material depending on its process history. Beta-amylase can only act where it can physically bind chain ends and align α-1,4 bonds in the active site.
Gelatinization increases accessibility by disrupting granule order and allowing water into the starch structure. Liquefaction reduces viscosity and creates additional chain ends. Milling increases surface area. Germination activates endogenous enzyme systems and modifies grain structure. Each of these changes can increase the practical opportunity for beta-amylase to release maltose.
The enzyme’s multiple-attack behavior makes accessibility even more important. If beta-amylase can remain associated with a chain and slide to successive cleavage positions, it can release several maltose units efficiently. If the chain is blocked, branched, aggregated, or inaccessible, that advantage is lost and the enzyme must search for another productive chain end [5].
This is also why beta-amylase is not best described as a universal “starch liquefier.” It is a targeted saccharification enzyme. Its strongest contribution appears after the process has created accessible non-reducing ends and when the desired sugar profile rewards maltose formation.
The main benefit of beta-amylase is controlled maltose enrichment. Instead of relying only on random starch fragmentation, the process can be directed toward a sugar that is highly relevant in brewing, distilling, cereal fermentation, and maltose syrup applications. The enzyme’s exo-action gives it a predictable product tendency that follows from its chain-end mechanism [1].
A second benefit is complementarity with other amylases. In alpha amylase beta amylase systems, alpha-amylase can open starch internally while beta-amylase converts the resulting chain ends into maltose. This helps explain why alpha and beta amylase are often discussed together in brewing and starch processing: they act on the same substrate family but produce different functional outcomes.
A third benefit is process interpretation. When maltose formation is lower than expected, the mechanism points to likely causes: insufficient starch accessibility, excessive heat exposure, branch-point limitation, or an enzyme mix that does not create enough suitable chain ends. This kind of mechanistic understanding is more useful than treating enzyme addition as a black box.
Finally, beta-amylase supports product differentiation where maltose matters. A maltose-rich wort, mash, hydrolysate, or syrup is chemically different from a glucose-rich or dextrin-rich one. Beta-amylase gives processors a specific way to move toward that profile, provided the substrate and process stage are appropriate.
Enzymes.bio supplies Beta-Amylase directly online by the 1 kg unit for commercial users who need a maltose-forming enzyme for starch conversion work. The product is purchased through the online store: the buyer places the order, pays online, and the order is processed and shipped. A Certificate of Analysis and Safety Data Sheet are included with the order.
For buyers searching “beta amylase enzyme for sale,” the key application fit is maltose-rich conversion of accessible starch. Beta-amylase is especially relevant in brewing, distilling, cereal processing, fermentation feed preparation, and high-maltose syrup workflows where the desired effect is chain-end release of maltose rather than broad, random starch thinning.
The science behind the enzyme is well established: beta-amylase acts from non-reducing ends, hydrolyzes accessible α-1,4 linkages, releases maltose, and stops at structural limits such as amylopectin branch points. Its best use is therefore as a targeted maltogenic enzyme within a starch-conversion process, not as a stand-alone solution for every starch-processing requirement.
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Beta-Amylase →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.