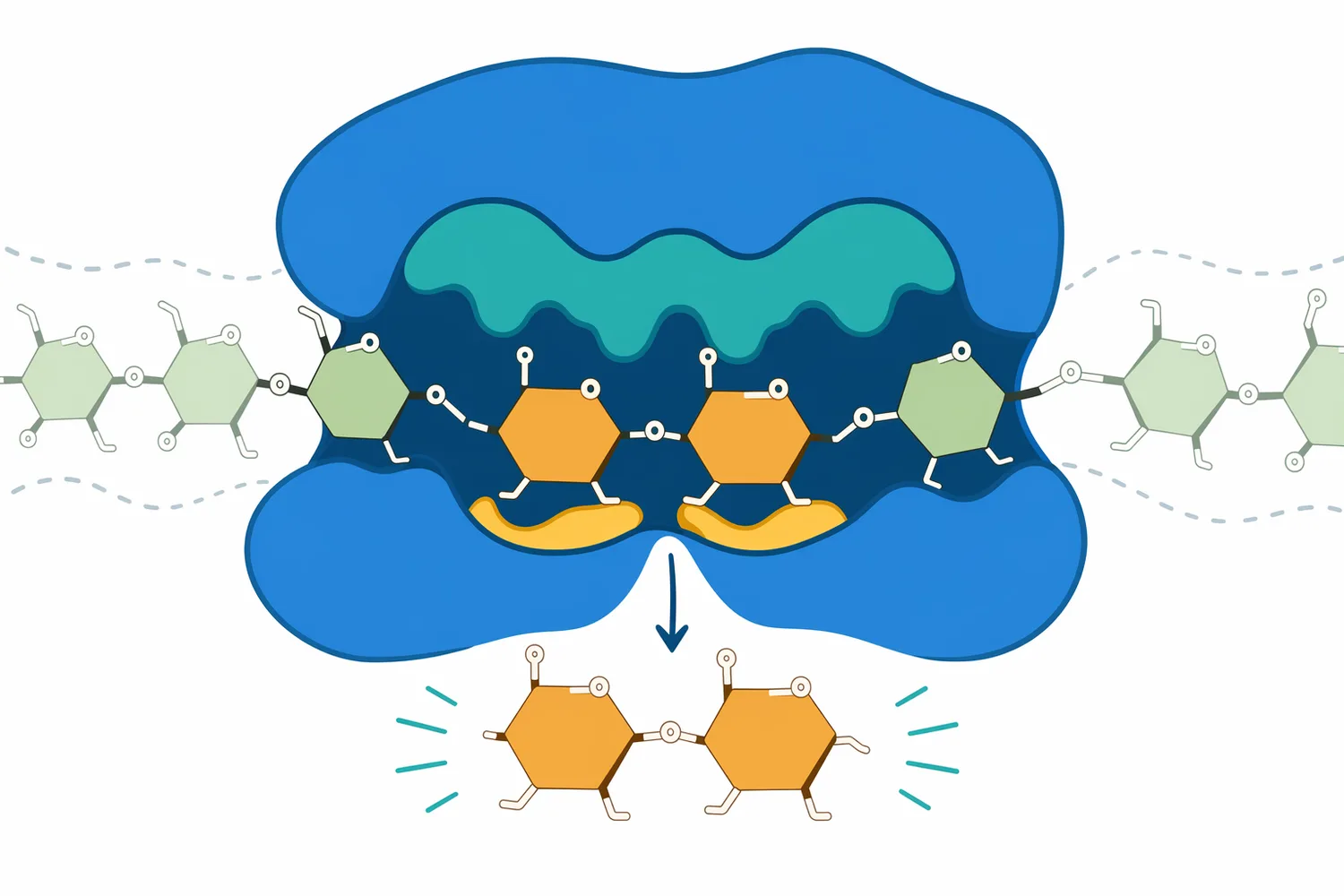
Beta-Amylase(β-アミラーゼ、beta amylase)は、デンプンやデンプン液化物の非還元末端から主にマルトースを切り出す糖化酵素で、高マルトースシロップ、ビール醸造、酢などの発酵前糖化で重要です。α-アミラーゼが内部切断で粘度を下げる酵素であるのに対し、β-アミラーゼは糖組成をマルトース側へ寄せる工程設計に向きます。分岐をもつアミロペクチンでは限界デキストリンが残るため、実務ではα-アミラーゼやプルラナーゼなどとの組み合わせが収率と糖組成を左右します[1]。
Beta-Amylaseは、デンプン、アミロース、アミロペクチン、マルトデキストリンなどのα-1,4-グルカン鎖に作用し、非還元末端からマルトース単位を順次遊離する酵素です。デンプン加工で使われるアミラーゼ群には、内部切断型のα-アミラーゼ、末端からグルコースを生成するグルコアミラーゼ、分岐を切るプルラナーゼなどがありますが、β-アミラーゼの特徴は「主生成物をマルトースに寄せる」点にあります。アミラーゼの微生物生産と産業応用に関する近年のレビューでも、アミラーゼ群は食品、発酵、デンプン加工などのバイオテクノロジー産業で中心的な酵素群として扱われています[2]。
β-アミラーゼの「β」は、生成されるマルトースのアノマー型に由来する名称であり、β-グルカンを分解する酵素という意味ではありません。この点は実務上も重要です。たとえば、麦芽、穀類、根菜由来デンプン、液化デンプンを扱う工程では「β-アミラーゼ=デンプンからマルトースを増やす酵素」と理解するのが適切で、β-グルカナーゼやセルラーゼとは基質も用途も異なります。植物由来原料では、ソバなどの穀類・擬穀類にもα-アミラーゼとβ-アミラーゼ活性が報告されており、デンプン貯蔵組織の糖化に両者が関与することが示されています[3]。
英語表記では beta-amylase とハイフンを入れる表記が一般的ですが、検索や調達文書では beta amylase と書かれることもあります。どちらも通常は同じ酵素群を指しますが、由来、安定性、適用工程、食品加工用途での位置づけは製品ごとに異なります。Enzymes.bioでは、Beta-AmylaseをB2B向けの酵素製品としてオンラインで提供しており、1kg単位で直接購入でき、注文時にCoAおよびSDSが併せて提供されます。
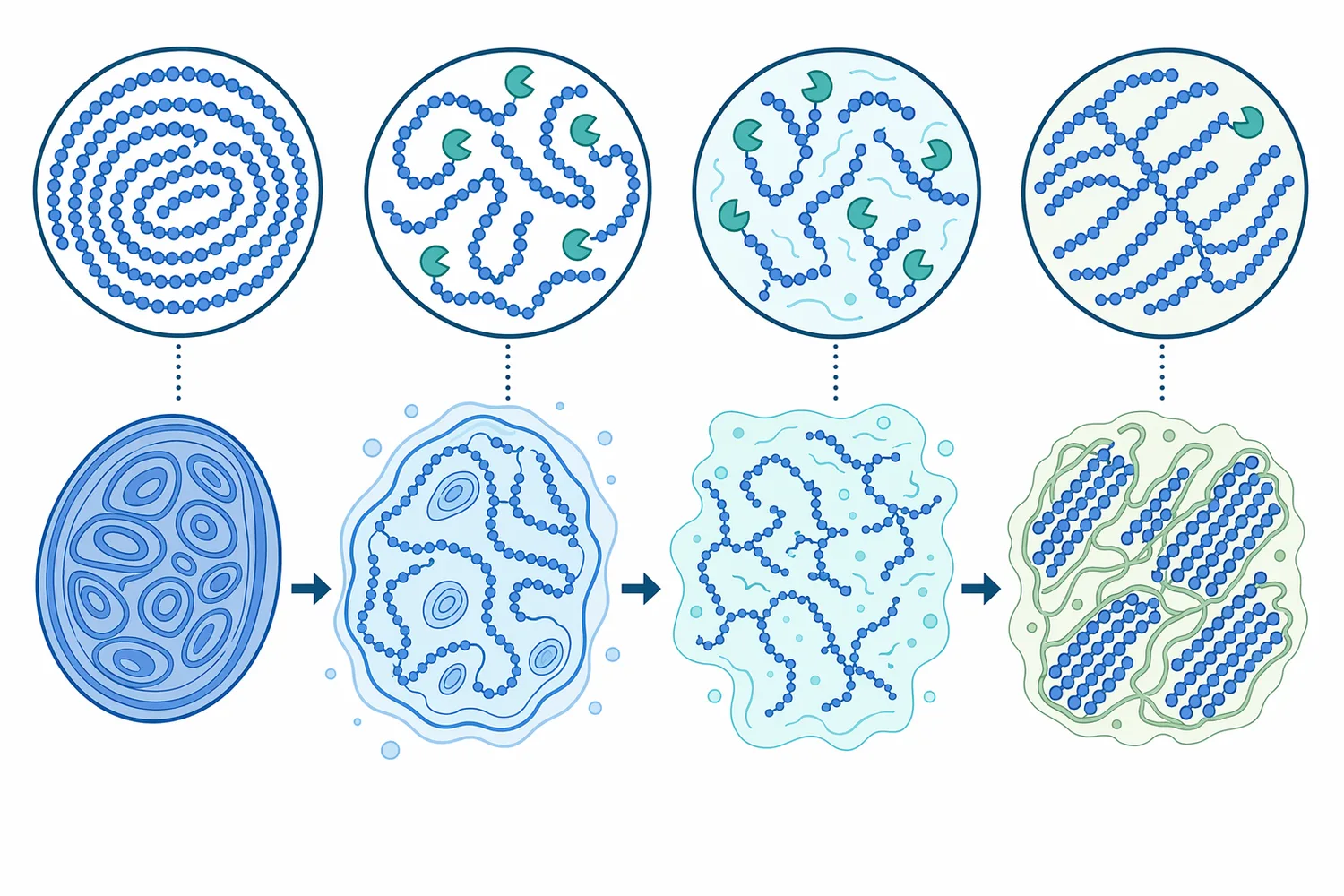
デンプンは、グルコースがα-1,4結合で直鎖状につながったアミロースと、α-1,4主鎖にα-1,6分岐をもつアミロペクチンから構成されます。Beta-Amylaseは主にα-1,4結合を標的としますが、鎖の途中をランダムに切るのではなく、非還元末端から2糖単位ずつ切り進みます。このため、基質鎖の末端数、液化によって生じた短鎖の分布、分岐点の密度が反応速度と最終糖組成を大きく左右します。サツマイモ由来β-アミラーゼの構造動態を扱った研究でも、酵素の立体構造と基質認識がβ-アミラーゼ機能を理解するうえで重要であることが示されています[4]。
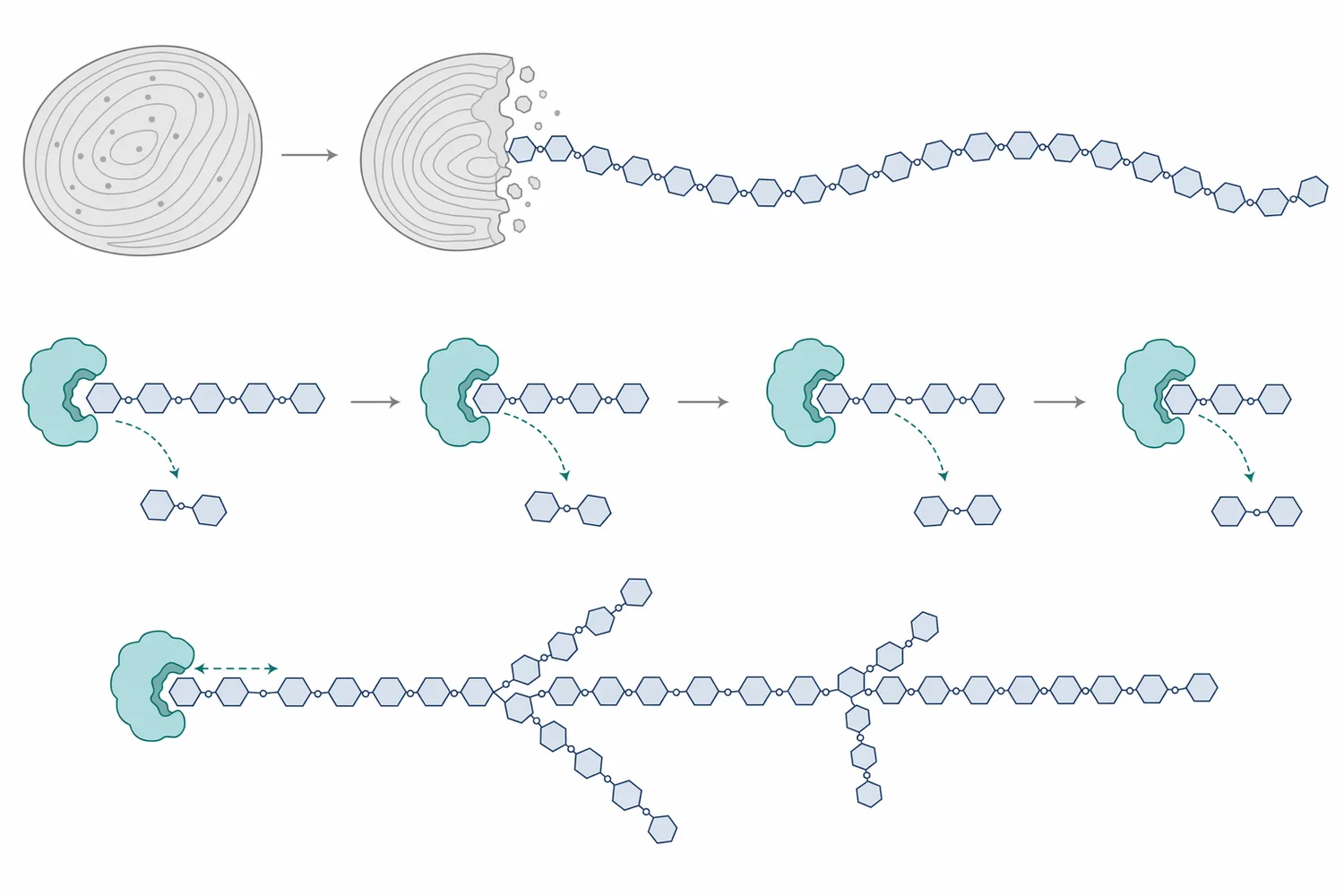
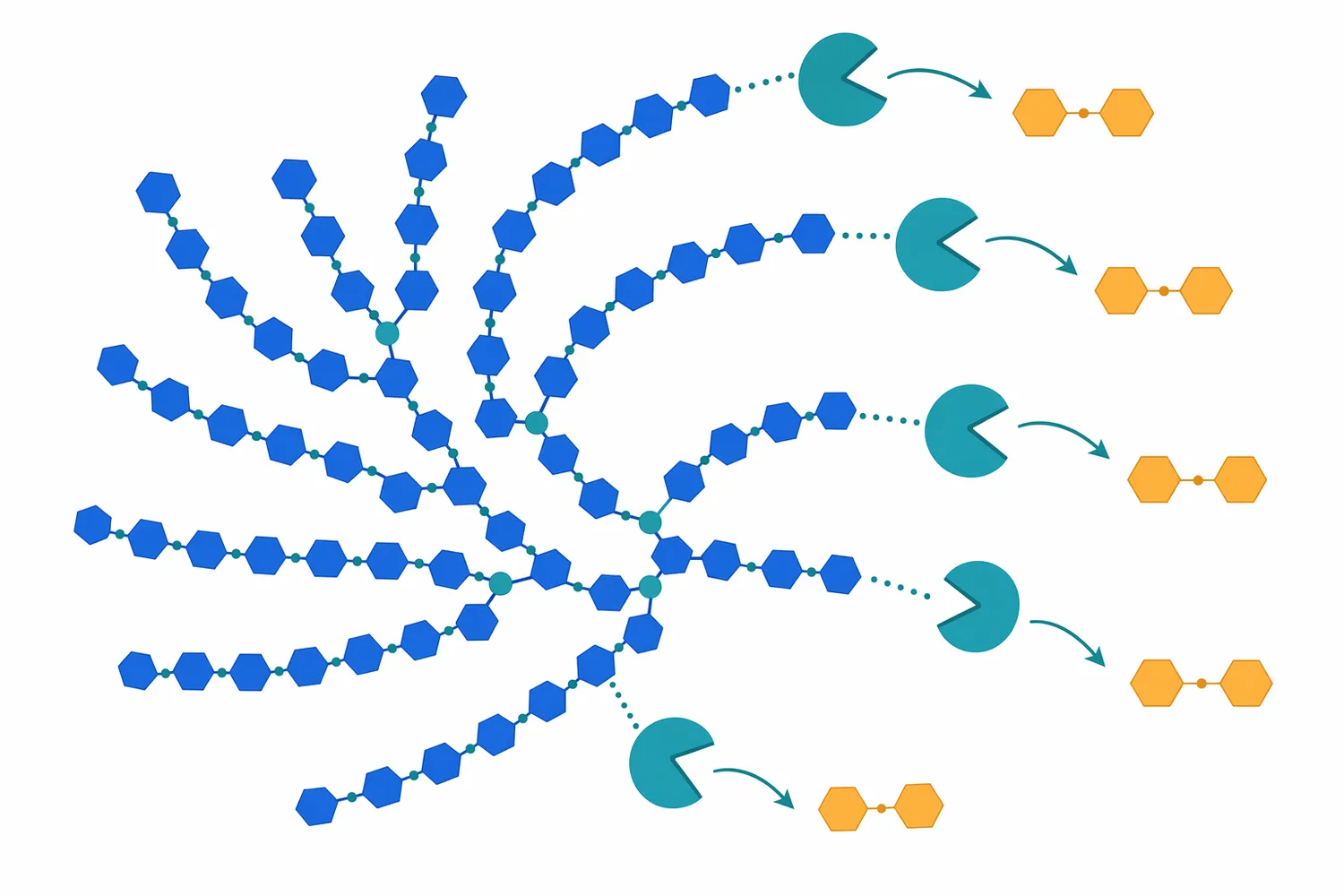
アミロースのような直鎖成分では、β-アミラーゼは比較的連続的にマルトースを生成できます。一方、アミロペクチンでは分岐点であるα-1,6結合の近傍まで進むと反応が止まり、β-リミットデキストリンと呼ばれる分岐残渣が残ります。これはβ-アミラーゼの欠点というより、酵素の作用様式から生じる必然的な制約です。そのため、高マルトース化をさらに進めたい場合には、分岐構造を減らす酵素、すなわちプルラナーゼなどの脱分岐酵素との工程設計が意味を持ちます。高濃度マルトデキストリン溶液を対象にした研究では、プルラナーゼ前処理がβ-アミラーゼ糖化時のマルトース収率を改善することが報告されています[1]。
この機序は、β-アミラーゼを「デンプンを完全にグルコースへ変える酵素」と誤解しないためにも重要です。グルコース主体の糖化を狙う場合はグルコアミラーゼが中心になりますが、マルトース主体の糖液を得たい場合はβ-アミラーゼの役割が大きくなります。目的が粘度低下なのか、発酵性糖の確保なのか、マルトース比率の上昇なのかによって、同じ「デンプン糖化」でも選ぶ酵素は変わります。α-アミラーゼとβ-アミラーゼの混合系では相乗作用に関する研究もあり、両酵素の組み合わせが単純な足し算ではない可能性が議論されています[5]。
デンプン加工では、複数の酵素名が混在しやすく、Beta-Amylaseの用途を正確に理解するには他酵素との違いを整理する必要があります。α-アミラーゼはデンプン鎖の内部を切って粘度を下げ、液化を進めます。β-アミラーゼは液化後または部分分解後の鎖末端からマルトースを生成します。グルコアミラーゼは末端からグルコースを生成し、プルラナーゼはα-1,6分岐を切ってβ-アミラーゼやグルコアミラーゼが作用しやすい直鎖を増やします。酵素触媒を工業製造に利用する際は、単一酵素の性質だけでなく、基質構造、反応順序、目的生成物に応じて組み合わせを設計することが重要です[6]。
| 酵素 | 主な作用位置 | 主生成物・工程効果 | Beta-Amylaseとの関係 |
|---|---|---|---|
| α-アミラーゼ | デンプン鎖内部のα-1,4結合 | 粘度低下、液化、短鎖デキストリン生成 | β-アミラーゼが作用できる末端を増やし得る |
| β-アミラーゼ | 非還元末端側のα-1,4結合 | 主にマルトース生成 | 高マルトース糖化の中心酵素 |
| グルコアミラーゼ | 非還元末端側 | 主にグルコース生成 | グルコース主体糖化では有利だが、マルトース主体設計とは目的が異なる |
| プルラナーゼ | α-1,6分岐結合 | 脱分岐、直鎖化 | β-アミラーゼの到達限界を緩和し、マルトース生成を支援し得る |
この比較から分かるように、β-アミラーゼは「液化酵素」ではなく「マルトース生成酵素」と捉えるのが実務的です。液化が不十分で基質粘度が高い場合、酵素が基質に均一に接触しにくく、反応が設計通りに進みにくくなります。一方、過度に別酵素で分解しすぎると、目的とするマルトース主体の糖組成から外れる可能性があります。特定マルトオリゴ糖の生成を狙う研究では、アミラーゼと分岐酵素などの組み合わせにより、生成物分布を制御する考え方が示されています[7]。
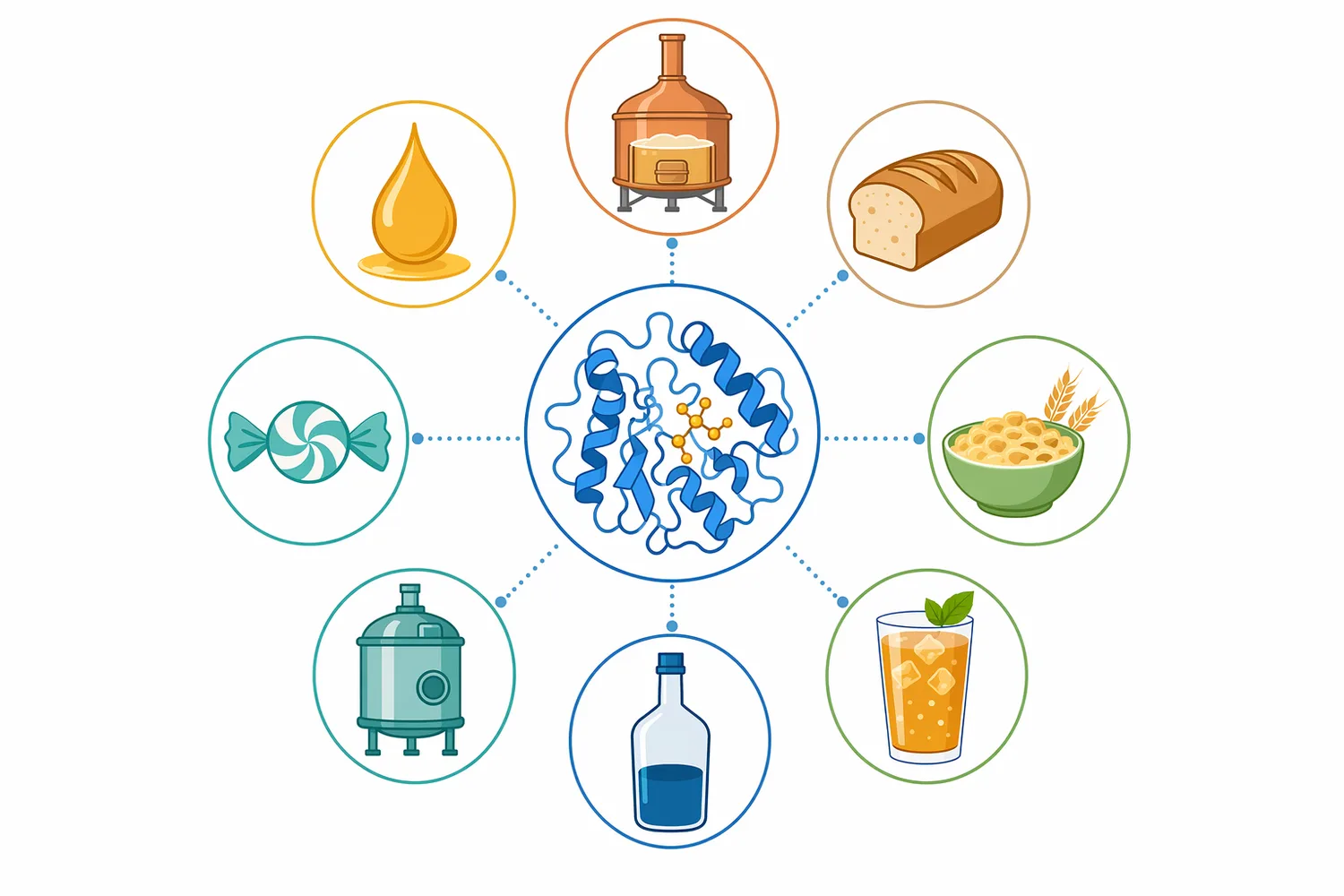
高マルトースシロップでは、甘味、発酵性、粘度、結晶化傾向、加熱時の反応性などを狙って、糖液中のマルトース比率を高めます。Beta-Amylaseは非還元末端からマルトースを生成するため、この用途に適した酵素です。特に、α-アミラーゼで液化されたデンプン、またはマルトデキストリンを出発基質とし、β-アミラーゼで糖化する設計では、液化の程度と分岐構造がマルトース生成に直結します。高濃度マルトデキストリンを対象にしたβ-アミラーゼ糖化研究では、脱分岐前処理によってマルトース収率が改善することが示されており、高マルトース工程で分岐構造が重要な制御点であることを裏付けます[1]。
高マルトースシロップは、グルコースシロップや高果糖シロップとは性質が異なります。マルトースは二糖であるため、同じ固形分でも浸透圧、甘味の立ち上がり、酵母による利用、加熱時の褐変挙動が異なります。したがって、β-アミラーゼの価値は単に「糖を増やす」ことではなく、糖組成をマルトース寄りに設計できる点にあります。Enzymes.bioのBeta-Amylaseは、食品加工・産業工程での利用を想定したB2B酵素製品として提供されており、高マルトースシロップを含むデンプン糖化用途に位置づけられます。
実務では、デンプン原料の種類も結果に影響します。トウモロコシ、タピオカ、小麦、米、馬鈴薯などはアミロースとアミロペクチンの比率、顆粒構造、リン酸基や脂質複合体の有無が異なります。β-アミラーゼは可溶化・糊化していないデンプン粒には十分作用しにくい場合があるため、前段の加熱、糊化、液化、せん断、固形分濃度の設計が重要になります。アミラーゼ生産と応用を扱うレビューでも、産業利用では酵素特性と基質条件の適合がプロセス性能を左右する要因として整理されています[2]。
ビール醸造では、麦芽由来デンプンを糖化して酵母が利用できる糖に変換します。麦汁中のマルトースは発酵性糖の主要成分の一つであり、β-アミラーゼはこのマルトース形成に関与します。麦芽には天然の酵素系が存在しますが、副原料の使用、原料ばらつき、糖化設計、発酵性の調整によっては、外部酵素の役割が検討されます。植物組織におけるα-アミラーゼとβ-アミラーゼの併存は、穀類・擬穀類原料の糖化を理解するうえでも参考になります[3]。
醸造工程で重要なのは、β-アミラーゼ単独の反応ではなく、温度休止、麦芽由来α-アミラーゼ、デキストリン生成、酵母の糖利用性との相互作用です。β-アミラーゼを働かせやすい条件を長く取りすぎると、発酵性が高まり、ボディ感が変化する可能性があります。一方、デキストリンを残す設計では、β-アミラーゼ作用を制限することもあります。α-アミラーゼとβ-アミラーゼの混合系に関する研究は、両酵素の相互作用が糖化プロファイルに影響し得ることを示しており、醸造のような複合酵素系では反応を一つの酵素だけで説明しない姿勢が必要です[5]。
酢やその他の発酵食品でも、デンプン質原料から発酵に使える糖を得る工程が品質と歩留まりに関わります。β-アミラーゼはマルトース供給を支える酵素であり、酵母や麹由来酵素、乳酸菌、酢酸菌などの代謝と組み合わさって最終製品の糖残存、酸生成、香味形成に影響します。ただし、発酵速度や風味は微生物、栄養成分、酸度、温度、発酵時間にも強く依存するため、β-アミラーゼの添加だけで最終品質を一義的に決めることはできません。バイオカタリシス全般の応用でも、酵素反応はプロセス条件と下流工程に組み込まれて初めて産業的価値を持つと整理されています[8]。
Beta-Amylaseの効果は、基質の末端数と分岐構造に大きく依存します。液化が十分に進んだデンプンでは、長鎖のままのデンプンに比べて酵素が作用できる末端が増えますが、同時に短すぎる糖鎖や分岐残渣が多いと、期待するマルトース生成が頭打ちになることがあります。したがって、α-アミラーゼによる液化はβ-アミラーゼ糖化の前提になり得ますが、液化の強さは目的糖組成と連動して決める必要があります。アミラーゼ関連研究では、酵素の組み合わせによって特定のマルトオリゴ糖や糖化プロファイルを制御する方向性が示されています[7]。
アミロペクチンが多い原料では、α-1,6分岐がβ-アミラーゼの進行を止めるため、脱分岐酵素の意味が大きくなります。プルラナーゼは分岐結合を切ることで、β-アミラーゼがさらに非還元末端からマルトースを生成できる直鎖領域を増やします。高濃度マルトデキストリン溶液での研究では、プルラナーゼ前処理がβ-アミラーゼ糖化におけるマルトース収率の改善につながることが示されており、この関係は高マルトースシロップ設計の機序説明として特に有用です[1]。
一方で、グルコアミラーゼとの併用は目的に注意が必要です。グルコアミラーゼはグルコースを増やすため、発酵性を高めたい場合には有効ですが、マルトース主体のシロップを狙う場合には糖組成がずれる可能性があります。β-アミラーゼの工程価値は「デンプンを分解する力」そのものではなく、「マルトースを中心とした糖組成を作りやすいこと」です。この違いを明確にしておくと、高マルトースシロップ、ビール醸造、酢発酵、発酵原料糖化などで酵素選択を誤りにくくなります。酵素触媒を製造工程に組み込む際は、生成物選択性とプロセス全体の物質収支を合わせて考える必要があります[6]。
β-アミラーゼは植物に多く見られる酵素として知られ、麦芽、サツマイモ、大豆などのデンプン貯蔵・動員に関わる酵素群として研究されてきました。サツマイモ由来β-アミラーゼの構造動態を解析した研究は、植物β-アミラーゼの立体構造、安定性、基質結合を理解する手がかりを提供しています[4]。植物由来酵素は食品加工との親和性が高い一方、工程条件への適合性や安定性は由来と製剤設計によって異なります。
微生物由来β-アミラーゼも研究対象です。好塩性Halobacillus sp. LY9による細胞外β-アミラーゼ生産の報告は、β-アミラーゼが植物だけでなく微生物からも得られること、また環境由来微生物が産業酵素探索の対象になり得ることを示しています[9]。ただし、研究で報告された特定菌株の性質を、そのまま市販製品全般またはEnzymes.bioが供給する製品の性質として読み替えることは適切ではありません。
アミラーゼ全体では、細菌、真菌、植物、海洋由来微生物など多様な供給源が探索されています。海綿関連微生物からのアミラーゼ生産を扱った研究や、温泉堆積物由来の耐熱性細菌に関する報告は、産業酵素候補がさまざまな環境から見いだされていることを示します[10]。この多様性は、酵素の温度安定性、pH耐性、塩耐性、基質特異性を広げる可能性がありますが、個別製品の性能は製品情報と実工程条件に基づいて扱う必要があります。
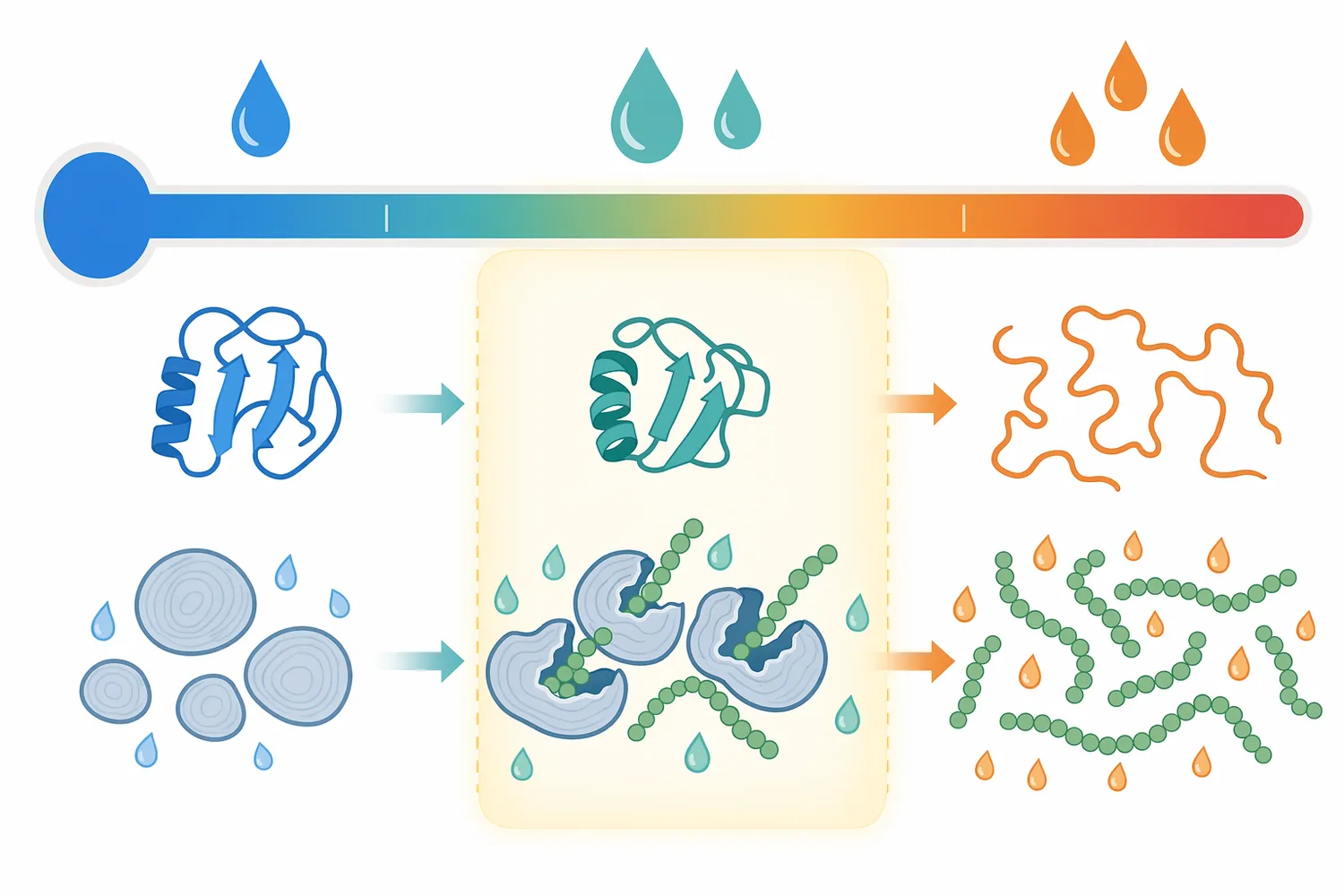
Beta-Amylaseはタンパク質触媒であり、温度、pH、水分活性、基質濃度、反応時間、金属イオン、阻害物質、せん断、熱履歴の影響を受けます。温度を上げると一般に反応速度は上がりますが、酵素の失活も進みやすくなります。pHが酵素の好適範囲から外れると、活性中心の電荷状態や立体構造が変化し、マルトース生成効率が低下します。酵素反応速度の理解には、基質濃度と反応速度の関係、失活、生成物蓄積などを同時に考える必要があり、酵素動力学教育でもこれらの動的要因が重要なテーマとして扱われています[11]。
高固形分の糖化では、基質濃度が高いほど製造効率は上がりますが、粘度上昇、混合不均一、生成物阻害、熱移動の不均一が起こりやすくなります。β-アミラーゼは末端から作用するため、基質鎖が物理的にアクセスしにくい状態では十分な効果を発揮しにくくなります。高濃度マルトデキストリンを対象にした研究でプルラナーゼ前処理が有効だったことは、単に酵素量を増やすよりも、基質構造と反応順序を整えることが重要であることを示しています[1]。
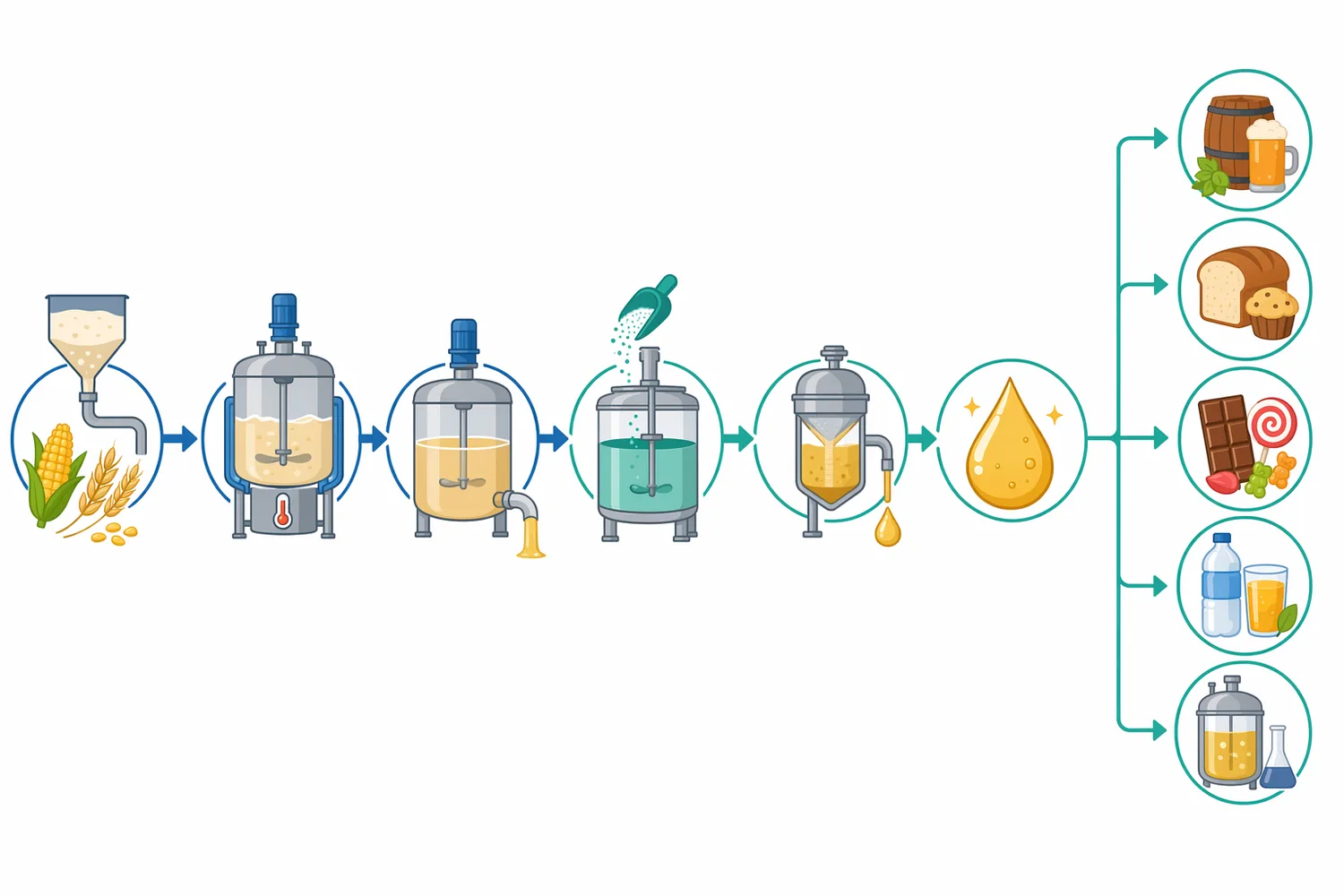
食品・発酵工程では、熱処理による酵素失活のタイミングも重要です。糖化後に酵素反応を止める必要がある場合、加熱やpH変化など工程内の既存操作を利用することがありますが、過度な加熱は糖液の色調や風味にも影響します。また、発酵工程に酵素が残る場合、発酵中も糖組成が変化し続ける可能性があります。バイオカタリシスの産業応用では、反応工程と後段処理を切り離さず、全体の品質設計として酵素を位置づけることが求められます[8]。
Enzymes.bioは酵素を供給するB2Bオンラインサプライヤーであり、製造業者や研究機関としてではなく、食品加工・産業用途向け酵素製品をオンラインで提供する立場です。Beta-Amylaseは1kg単位でオンライン直接販売され、注文時にはCoAおよびSDSが併せて提供されます。製品は工程投入用の酵素製剤として扱うべきであり、家庭用の直接摂取品、医薬品、サプリメントとして位置づけるものではありません。
この位置づけを明確にする理由は、β-アミラーゼの性能が工程条件に依存するためです。Enzymes.bioが供給するBeta-Amylaseは、高マルトースシロップ、ビール醸造、酢発酵など、マルトース生成が意味を持つ用途で検討される酵素です。ただし、最終的なマルトース比率、発酵速度、香味、歩留まり、粘度は、原料、液化条件、脱分岐の有無、反応時間、温度、pH、固形分濃度、後段発酵の微生物条件に左右されます。アミラーゼの産業応用に関するレビューでも、酵素の利用価値は基質とプロセス設計の適合によって決まることが示されています[2]。
また、β-アミラーゼは単独で万能なデンプン糖化酵素ではありません。高マルトース化には向きますが、完全なグルコース化や強い液化を主目的とする場合は、他の酵素が中心になります。逆に、グルコースを増やしすぎたくない、マルトース主体の糖液を得たい、発酵性とデキストリン残存のバランスを取りたい工程では、β-アミラーゼの選択性が価値になります。α-アミラーゼとβ-アミラーゼの相乗作用に関する研究は、複数酵素を組み合わせた糖化設計が、単一酵素では得にくい糖組成制御につながる可能性を示しています[5]。
Beta-Amylaseはタンパク質性の酵素であり、粉末製剤では粉じんの吸入、皮膚や眼への接触、作業環境中の飛散に注意が必要です。酵素一般に、反復吸入や接触によって感作が起こる可能性があるため、食品加工用の酵素であっても、作業者保護を前提に扱う必要があります。Enzymes.bioでは注文時にSDSが提供されるため、実際の取り扱い、保管、廃棄、漏えい時対応は当該SDSに従うことが基本です。
保管では、酵素の失活を避けるため、湿気、過度な熱、直射日光、開封後の吸湿を抑えることが重要です。酵素粉末は水分を吸うと凝集しやすく、局所的な活性低下や投入時の分散不良につながる場合があります。食品・発酵工程で使用する場合でも、衛生的な保管、交差汚染防止、適切なラベル管理は通常の工程管理の一部として扱われます。酵素触媒の産業利用では、反応効率だけでなく、安定性、保管性、操作性も実装時の重要な要素です[8]。
Beta-Amylaseについて比較的確実に言えるのは、デンプン由来基質から主にマルトースを生成する酵素であり、高マルトース化、醸造糖化、発酵前糖化に適した選択性を持つという点です。また、アミロペクチンの分岐が反応限界を作るため、プルラナーゼなどの脱分岐酵素と組み合わせるとマルトース生成が改善し得ることも、研究で支持されています[1]。
一方で、特定原料での最終マルトース収率、発酵速度、アルコール収率、酢酸生成速度、最終製品の香味、色調、保存安定性については、β-アミラーゼだけで断定できません。たとえば、同じbeta-amylaseを使っても、液化の程度、原料中のアミロペクチン比率、pH、温度、固形分、反応時間、併用酵素、微生物の糖利用性が異なれば、最終結果は変わります。酵素反応は高い選択性を持つ一方、工程全体の条件に組み込まれて初めて目的性能を発揮するためです[6]。
したがって、Beta-Amylaseは「マルトース生成を支える実用酵素」として理解するのが最も正確です。デンプンを完全に単糖化する酵素でも、液化を主目的とする酵素でもなく、非還元末端からマルトースを生み出す選択性によって価値を持ちます。高マルトースシロップ、ビール醸造、酢発酵、発酵原料糖化など、マルトースが工程品質や発酵性に関わる場面で、β-アミラーゼは中心的な選択肢になります。Enzymes.bioのBeta-Amylaseは、1kg単位でオンライン購入できるB2B向け酵素製品として、このような食品加工・産業用途での利用を想定して提供されています。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Beta-Amylaseを購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。