Beta-Amylase는 전분·글리코겐 같은 α-글루칸의 비환원 말단에서 maltose 단위를 순차적으로 방출하는 exo형 전분 분해 효소입니다. α-amylase가 전분 사슬 내부를 절단해 dextrin과 다양한 올리고당을 만드는 데 비해, beta-amylase function의 핵심은 maltose 비율을 높이는 방향으로 당 조성을 설계하는 것입니다 [1].
주요 응용은 고 maltose 시럽, 맥아·곡물 mashing, 발효 가능한 당 조성 조절, 전분 구조 연구입니다. 다만 β-amylase는 α-1,6 가지 결합을 직접 제거하는 효소가 아니므로 amylopectin이 많은 전분에서는 β-limit dextrin이 남을 수 있고, 실제 전환율은 전분 상태·열 이력·다른 amylolytic enzyme과의 조합에 크게 좌우됩니다 [2].
Beta-Amylase, 또는 beta amylase는 전분 사슬의 비환원 말단(non-reducing end)에서 α-1,4 글리코시드 결합을 단계적으로 가수분해하여 maltose를 생성하는 효소입니다. 전분을 “긴 glucose 사슬”로 보면, beta-amylase mechanism은 사슬 중간을 무작위로 자르는 것이 아니라 끝에서 두 개의 glucose 단위씩 떼어내는 방식에 가깝습니다. 이 작용 때문에 β-amylase는 maltose syrup, 맥아 당화, 곡물 발효에서 “maltose-rich sugar profile”을 만들고자 할 때 기술적으로 중요합니다 [3].
전분은 직선형에 가까운 amylose와 가지가 많은 amylopectin으로 구성됩니다. β-amylase는 α-1,4 결합을 따라 진행할 수 있지만, amylopectin의 α-1,6 branch point를 만나면 더 이상 같은 방식으로 진행하지 못합니다. 그래서 β-amylase 단독 처리 후에는 branch point 주변 구조가 남아 β-limit dextrin이 형성될 수 있으며, 이 현상은 waxy corn starch에서 β-limit dextrin을 준비하고 특성을 검토한 연구에서도 핵심 개념으로 다루어졌습니다 [2].
효소 분류와 산업 문맥에서 beta-amylase는 “전분을 당으로 바꾸는 amylase”라는 큰 범주에 속하지만, 기능은 α-amylase나 glucoamylase와 다릅니다. amylase류는 식품, 전분 가공, 발효, 사료, 세정, 바이오공정 등에서 널리 응용되는 효소군으로 정리되지만, 실제 제품 설계에서는 “어떤 결합을 어디서 자르는가”가 최종 당 조성과 물성에 직접 연결됩니다 [4].
Beta-amylase function을 한 문장으로 요약하면 전분 유래 α-글루칸을 maltose 중심의 당 조성으로 전환하는 것입니다. maltose는 glucose 두 분자가 연결된 이당류이며, 효모 발효, 감미 프로파일, Maillard 반응 가능성, 시럽 점도와 같은 여러 품질 변수에 영향을 줄 수 있습니다. maltose syrup 생산 공정에서는 전분 액화 후 당화 단계에서 β-amylase가 maltose 생성 방향을 강화하는 효소로 검토됩니다 [3].
고 maltose 시럽에서는 단순히 전분 분해율만 높은 것이 목적이 아닙니다. α-amylase만으로 전분을 처리하면 내부 절단에 의해 다양한 길이의 dextrin과 oligosaccharide가 생기고, glucoamylase를 강하게 쓰면 glucose 비율이 커질 수 있습니다. 반면 beta-amylase는 maltose를 반복적으로 방출하므로, 최종 당 조성을 maltose 쪽으로 설계하려는 식품·음료·발효 공정에 적합한 역할을 갖습니다 [1].
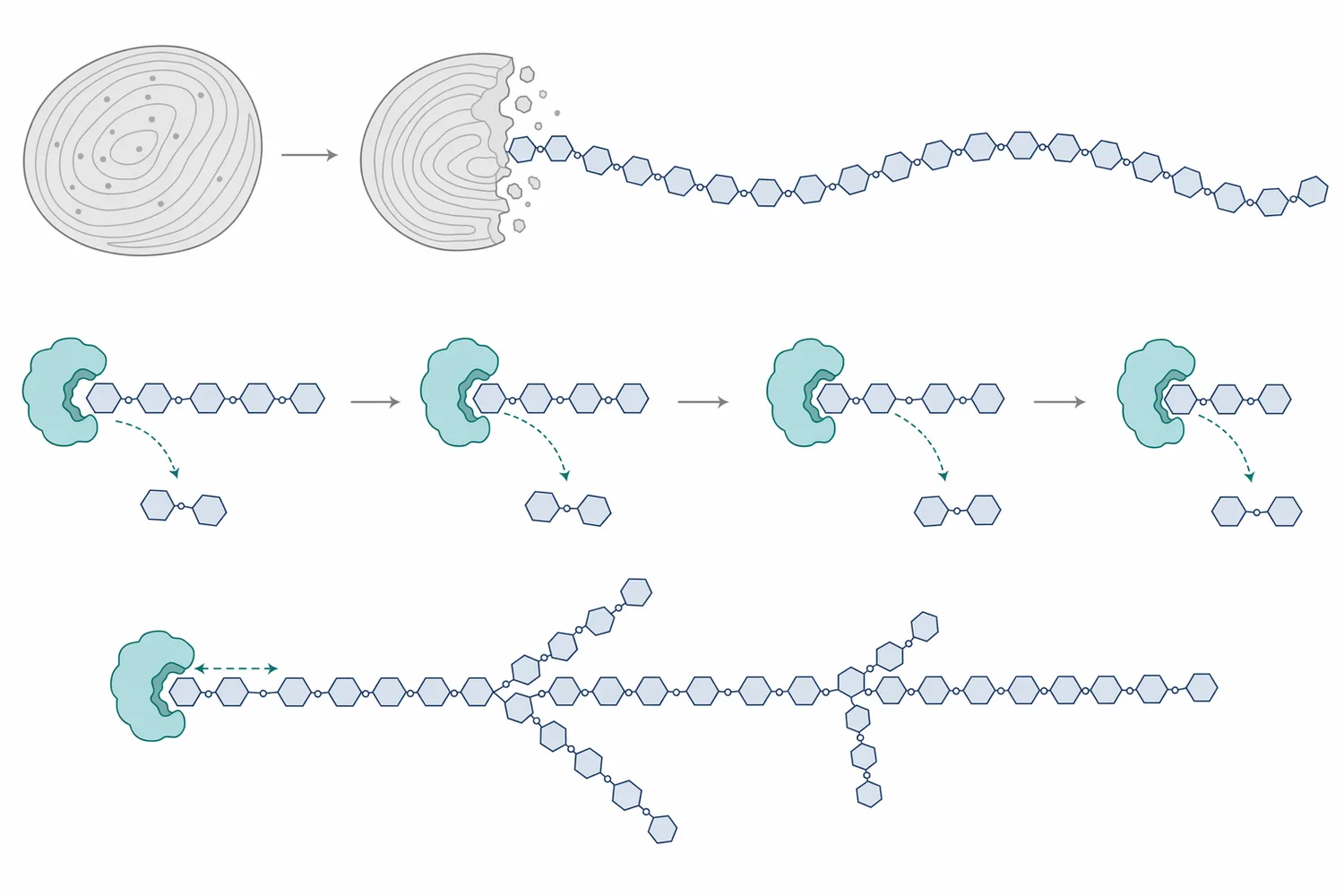
β-amylase의 장점은 “특정 당을 선택적으로 많이 만들 수 있다”는 점이지만, 한계도 같은 지점에서 나옵니다. 효소가 사슬 끝에서만 진행한다는 것은 전분 내부 접근성, 사슬 길이, 가지 구조, 전처리 상태가 모두 반응성에 영향을 준다는 뜻입니다. native starch granule의 가수분해가 입자 표면적과 관련된다는 연구는, 전분을 단순한 화학식이 아니라 표면·결정성·입도·팽윤 상태를 가진 물리적 기질로 보아야 함을 보여줍니다 [5].
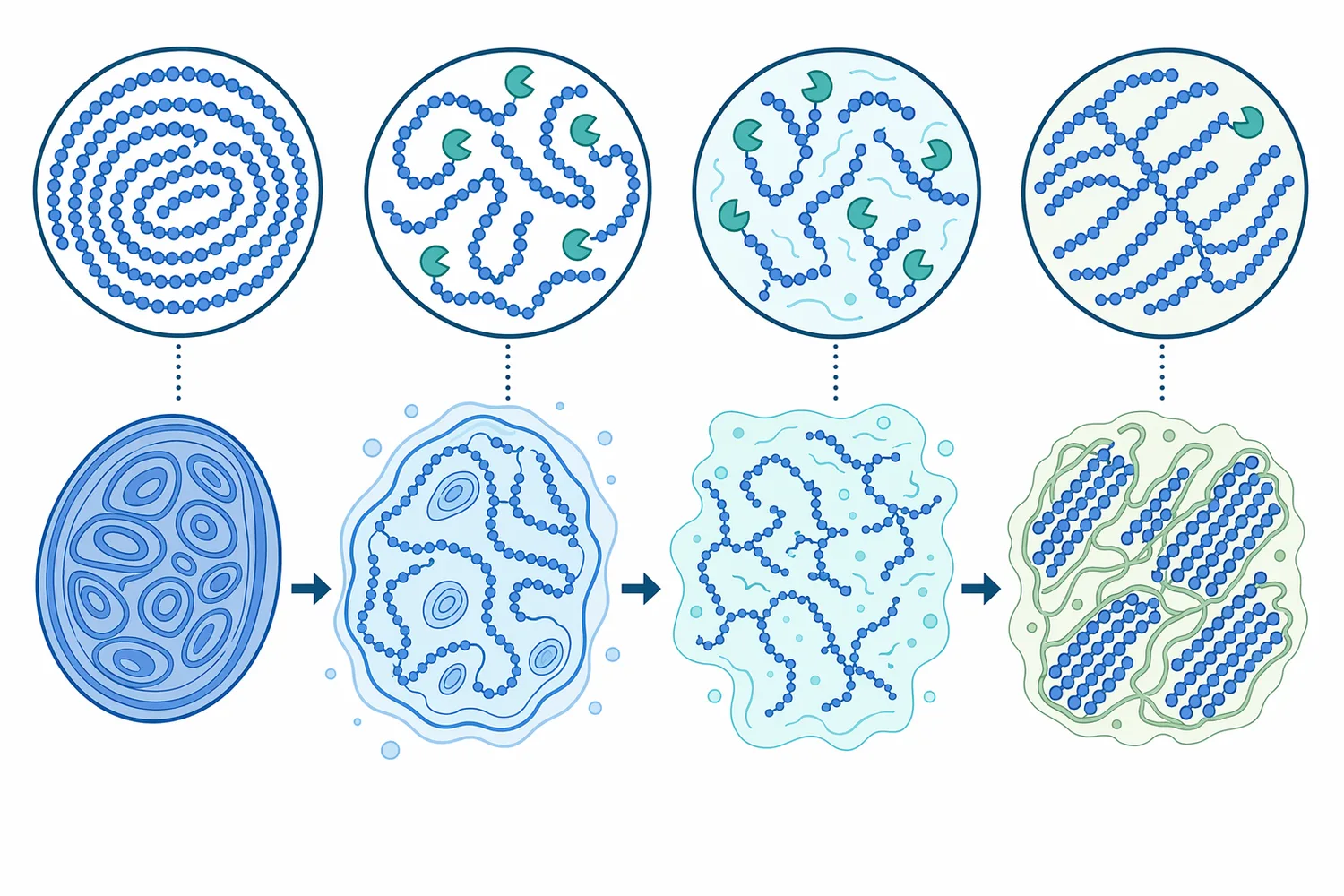
Beta-amylase mechanism에서 가장 중요한 세 요소는 비환원 말단, α-1,4 결합, branch point 정지입니다. 효소는 기질 사슬의 비환원 말단을 인식해 두 glucose 단위에 해당하는 maltose를 방출하고, 같은 방향으로 반복 작용합니다. 전분 사슬에 접근 가능한 비환원 말단이 많을수록 β-amylase가 작용할 수 있는 출발점도 늘어나며, 이 점이 α-amylase와의 조합에서 중요한 이유가 됩니다 [6].
α-amylase가 먼저 전분 내부를 절단하면 dextrin이 생성되고, 이 과정에서 새로운 사슬 말단이 증가합니다. 그런 다음 beta-amylase는 늘어난 비환원 말단에서 maltose를 방출할 수 있습니다. 즉 “alpha amylase beta amylase” 조합은 단순 병렬 사용이 아니라, α-amylase가 구조적으로 접근 가능한 기질을 만들고 β-amylase가 maltose를 축적하는 기능적 분업으로 해석할 수 있습니다 [6].
그러나 amylopectin의 α-1,6 가지 결합은 β-amylase가 직접 광범위하게 제거하는 대상이 아닙니다. β-amylase가 가지점 근처까지 진행하면 남는 구조가 β-limit dextrin이며, 이는 전분 구조 분석에서 오히려 유용한 정보가 됩니다. waxy corn starch에서 준비된 β-limit dextrin 연구는 β-amylase가 어디까지 작용하고 어디에서 멈추는지를 통해 amylopectin 구조와 잔류 dextrin 특성을 이해할 수 있음을 보여줍니다 [2].
“alpha amylase vs beta amylase”, “alpha vs beta amylase”, “difference between alpha and beta amylase”는 전분 공정을 설계할 때 가장 자주 등장하는 비교입니다. α-amylase는 endo형으로 전분 사슬 내부의 α-1,4 결합을 절단하여 점도를 빠르게 낮추고 dextrin을 생성하는 데 유리합니다. 반대로 β-amylase는 exo형으로 비환원 말단에서 maltose를 순차적으로 방출하므로, maltose 수율과 발효성 당 조성을 조절하는 쪽에서 의미가 큽니다 [7].
| 비교 항목 | α-Amylase | β-Amylase |
|---|---|---|
| 기본 작용 방식 | 전분 사슬 내부를 절단하는 endo형 | 비환원 말단에서 순차 절단하는 exo형 |
| 주요 생성물 경향 | dextrin, maltose, maltotriose 등 혼합 올리고당 | maltose 중심 |
| 공정상 대표 역할 | 액화, 점도 저감, 사슬 절단 | 당화, maltose 비율 강화 |
| 가지 구조 대응 | α-1,6 결합 자체를 전문적으로 제거하지는 않음 | α-1,6 branch point에서 진행 제한 |
| 조합 의미 | β-amylase가 작용할 말단을 늘릴 수 있음 | α-amylase가 만든 말단에서 maltose 방출 |
| 관련 검색어 맥락 | alpha-amylase beta amylase, alpha beta amylase | beta-amylase activity, beta amylase mechanism |
맥아 mashing 연구에서는 α-amylase, β-amylase, limit dextrinase가 각각 다른 방식으로 전분 분해에 기여한다고 모델링되었습니다. 이 조합에서 α-amylase는 사슬 내부 절단으로 dextrin 구조를 만들고, β-amylase는 maltose를 형성하며, limit dextrinase는 가지 결합 주변의 제한을 완화하는 역할로 이해됩니다 [6].
따라서 “beta amylase vs alpha amylase”의 답은 어느 효소가 더 좋으냐가 아니라, 목표가 무엇이냐에 따라 달라집니다. 점도 저감과 액화를 우선한다면 α-amylase의 역할이 커지고, maltose 중심 당화를 원한다면 β-amylase의 의미가 커집니다. 전분을 발효용 당으로 전환할 때는 두 효소의 순서와 열 이력, 원료 전분의 gelatinization 특성이 함께 작용합니다 [1].
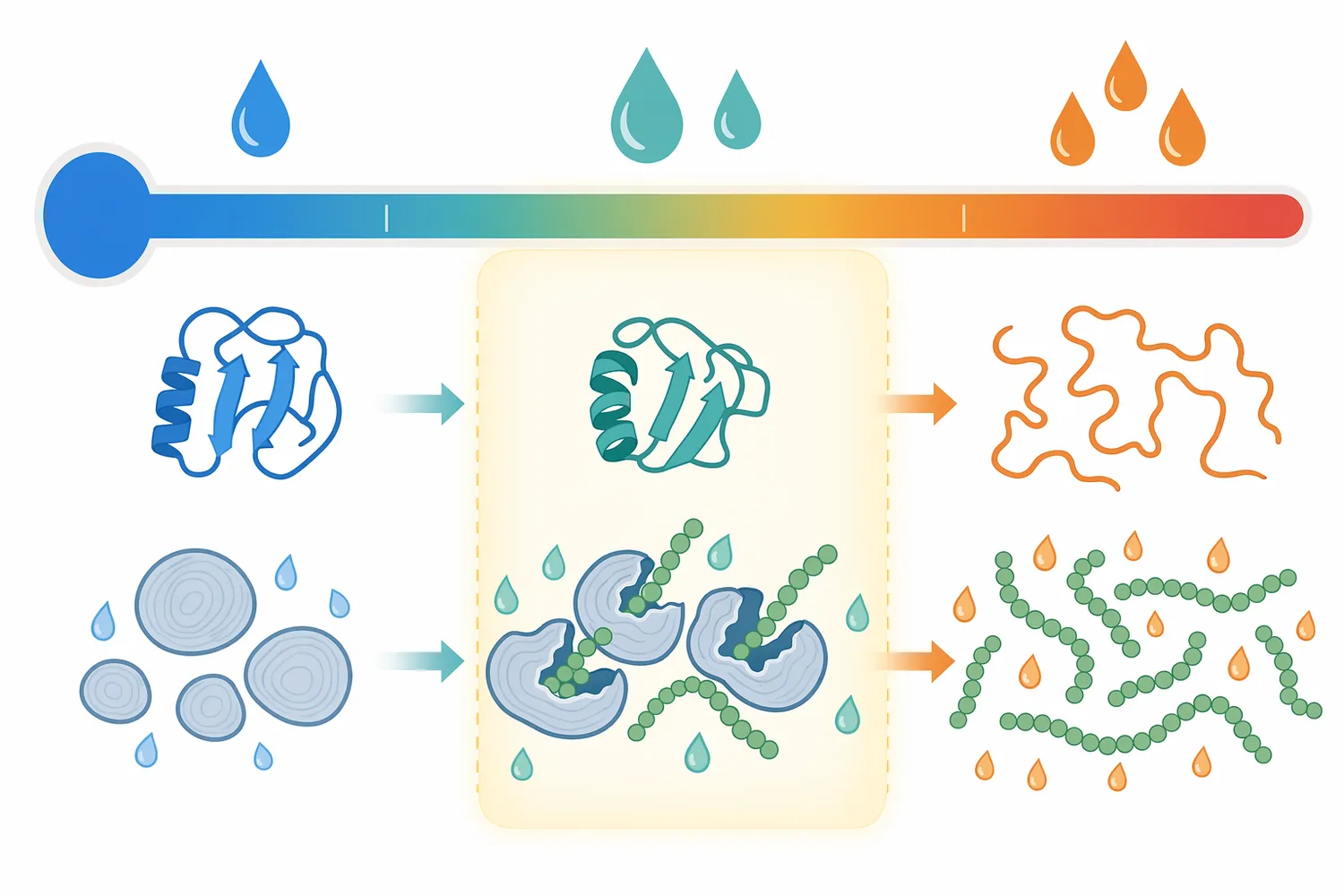
Beta-amylase activity는 효소 자체의 특성만으로 결정되지 않습니다. 같은 전분이라도 생전분 granule 상태인지, 부분 팽윤 상태인지, gelatinized starch인지, α-amylase로 액화된 dextrin 상태인지에 따라 효소가 접근할 수 있는 사슬 말단의 수와 노출 정도가 달라집니다. native starch granule 연구에서는 granule surface area가 amylolytic hydrolysis에 영향을 준다는 점이 강조되었고, 이는 전분 입자의 물리적 표면이 효소 반응을 제한할 수 있음을 의미합니다 [5].
전분 granule은 결정성 영역과 비결정성 영역이 섞인 입자 구조를 가지며, 물과 열을 받으면 팽윤과 gelatinization을 거쳐 효소 접근성이 증가할 수 있습니다. 그러나 과도한 열 이력은 효소 안정성에는 불리할 수 있고, 반대로 충분한 열 처리가 없으면 전분 사슬이 효소에 충분히 노출되지 않을 수 있습니다. 이 균형 때문에 실제 공정에서 beta-amylase temperature range는 단순한 숫자 하나가 아니라 기질 전처리, pH, 시간, 효소 조합이 함께 정의하는 운전 영역으로 이해해야 합니다 [3].
“alpha and beta amylase temperature range”를 비교할 때도 주의가 필요합니다. 일부 α-amylase는 액화 단계의 높은 온도 조건에 맞춰 쓰이고, β-amylase는 maltose 형성에 유리한 당화·mashing 조건에서 중요하게 작용하는 경우가 많습니다. 그러나 효소의 원천, 제형, 공정 pH, 전분 농도에 따라 안정성과 반응성은 달라지므로, 문헌상 모델은 특정 mashing 조건에서의 상대적 기여를 해석하는 근거로 보는 것이 타당합니다 [1].
맥아 mashing에서 β-amylase는 fermentable sugar 형성의 핵심 효소 중 하나입니다. 보리 맥아에는 전분 분해에 관여하는 여러 효소가 존재하며, mashing 중 온도와 시간에 따라 α-amylase, β-amylase, limit dextrinase의 상대적 기여가 달라집니다. 모델링 연구는 이 세 효소가 전분 분해 과정에서 서로 다른 위치와 기능을 맡는다는 점을 정량적으로 설명하려고 했습니다 [6].
맥주나 곡물 발효 공정에서 maltose는 효모가 이용할 수 있는 주요 당 중 하나입니다. β-amylase가 충분히 작용하면 maltose-rich wort 또는 당화액 형성에 기여할 수 있지만, 이것이 자동으로 발효 수율 향상을 보장한다는 뜻은 아닙니다. 수수 곡물의 에탄올 생산 연구에서는 효소 가수분해와 hydrolysate fermentability가 함께 평가되었고, 원료 특성과 가수분해 조건이 발효 결과에 영향을 주는 것으로 다루어졌습니다 [8].
따라서 β-amylase는 “발효를 무조건 높이는 효소”라기보다, 발효 가능한 당 조성을 maltose 중심으로 설계하는 도구로 보는 편이 정확합니다. 실제 발효 성능은 효모 균주가 maltose를 얼마나 효율적으로 수송·분해하는지, 잔류 dextrin 비율이 얼마나 되는지, 당화액의 질소원과 미량 성분이 충분한지, 삼투압과 pH가 적절한지에 의해 함께 결정됩니다 [8].
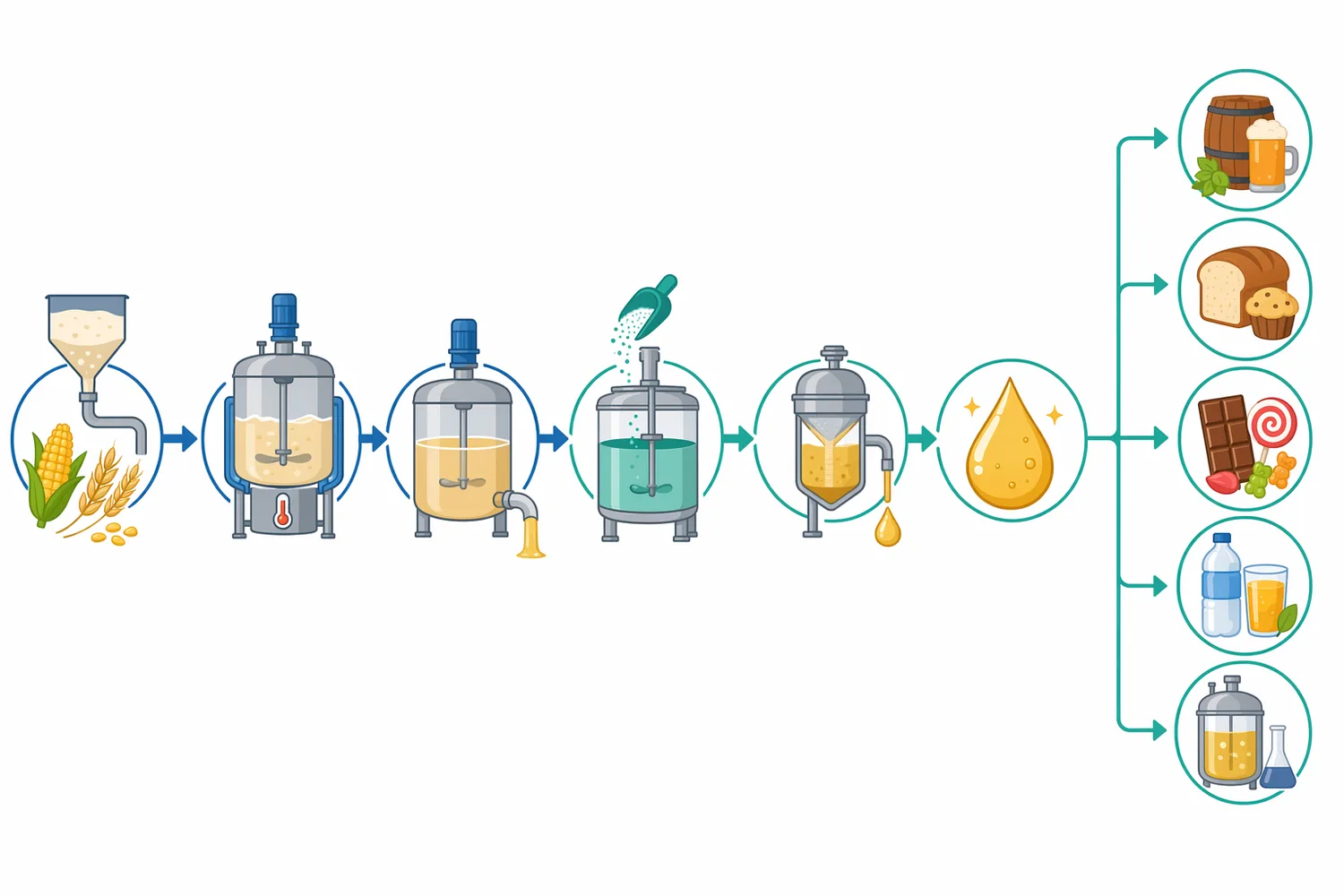
고 maltose 시럽은 식품 제조에서 감미, 점도, 갈변 반응, 발효성, 수분활성 조절과 연결될 수 있는 소재입니다. maltose syrup 생산 연구에서는 전분 전환 공정의 반응 조건과 공정 제어 변수가 최종 당 조성에 영향을 준다는 점이 다루어졌고, β-amylase는 maltose 생성을 강화하는 효소로 중요한 위치를 차지합니다 [3].
전형적인 전분 전환에서는 먼저 전분을 물리적으로 분산·가열하고, α-amylase 등으로 액화하여 점도를 낮춘 뒤, β-amylase가 접근 가능한 사슬 말단에서 maltose를 방출하도록 설계할 수 있습니다. 여기에 pullulanase 같은 debranching enzyme을 함께 쓰면 amylopectin 가지 구조로 인한 β-limit dextrin 잔류를 줄이고 maltose 형성 가능성을 높일 수 있습니다. β-amylase와 pullulanase의 고정화 two-enzyme system을 이용해 전분을 maltose로 전환하려는 연구는 이러한 조합 전략의 산업적 의미를 잘 보여줍니다 [9].
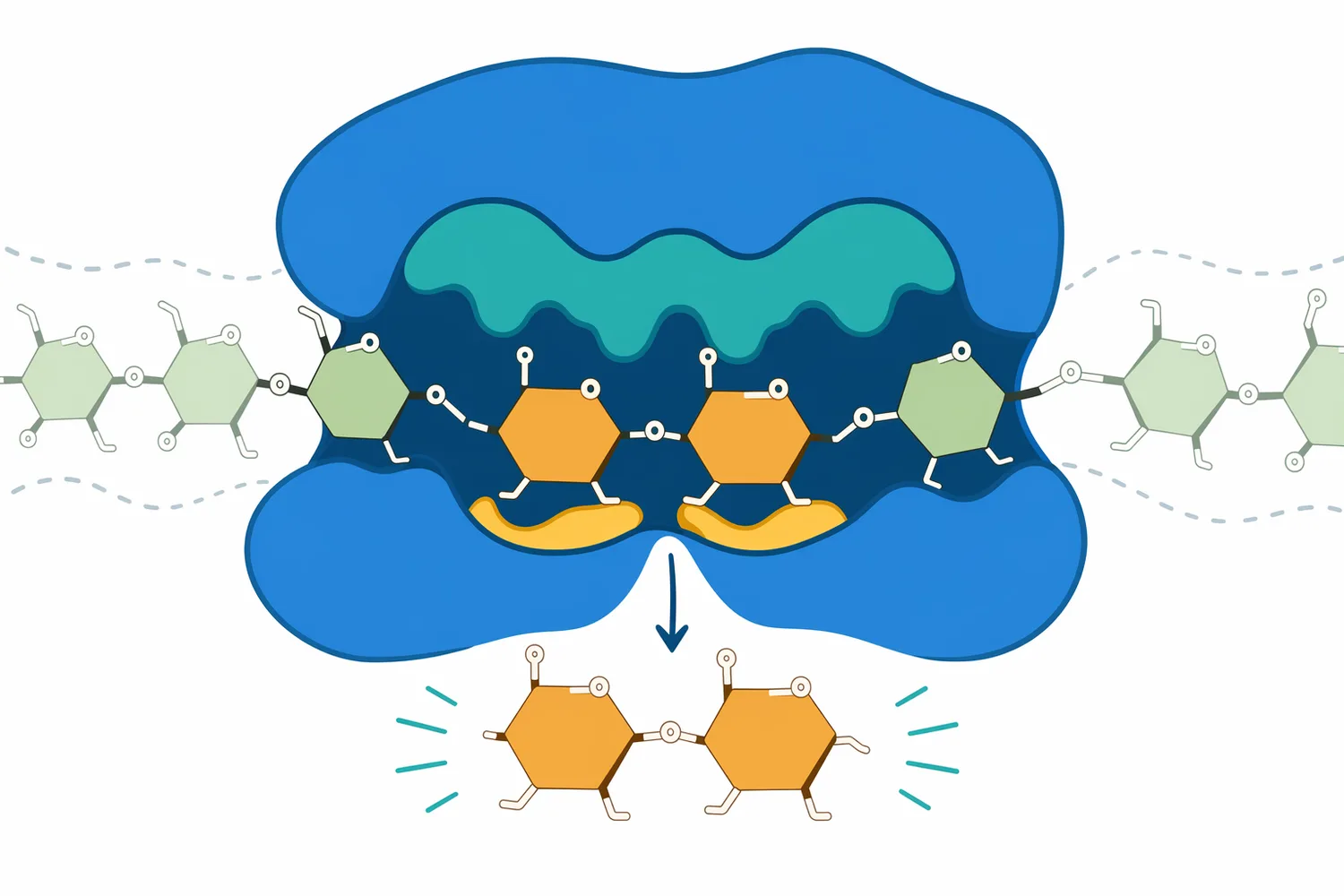
다만 고 maltose 시럽의 품질은 β-amylase만으로 결정되지 않습니다. 전분 원료가 옥수수인지, 타피오카인지, 감자인지, 곡물 전분인지에 따라 amylose/amylopectin 비율, granule 크기, gelatinization 거동이 달라지고, 이는 같은 효소 조합에서도 다른 당화 곡선을 만들 수 있습니다. native starch granule의 표면적이 가수분해에 영향을 준다는 연구는 원료 물성이 효소 반응의 중요한 독립 변수임을 뒷받침합니다 [5].
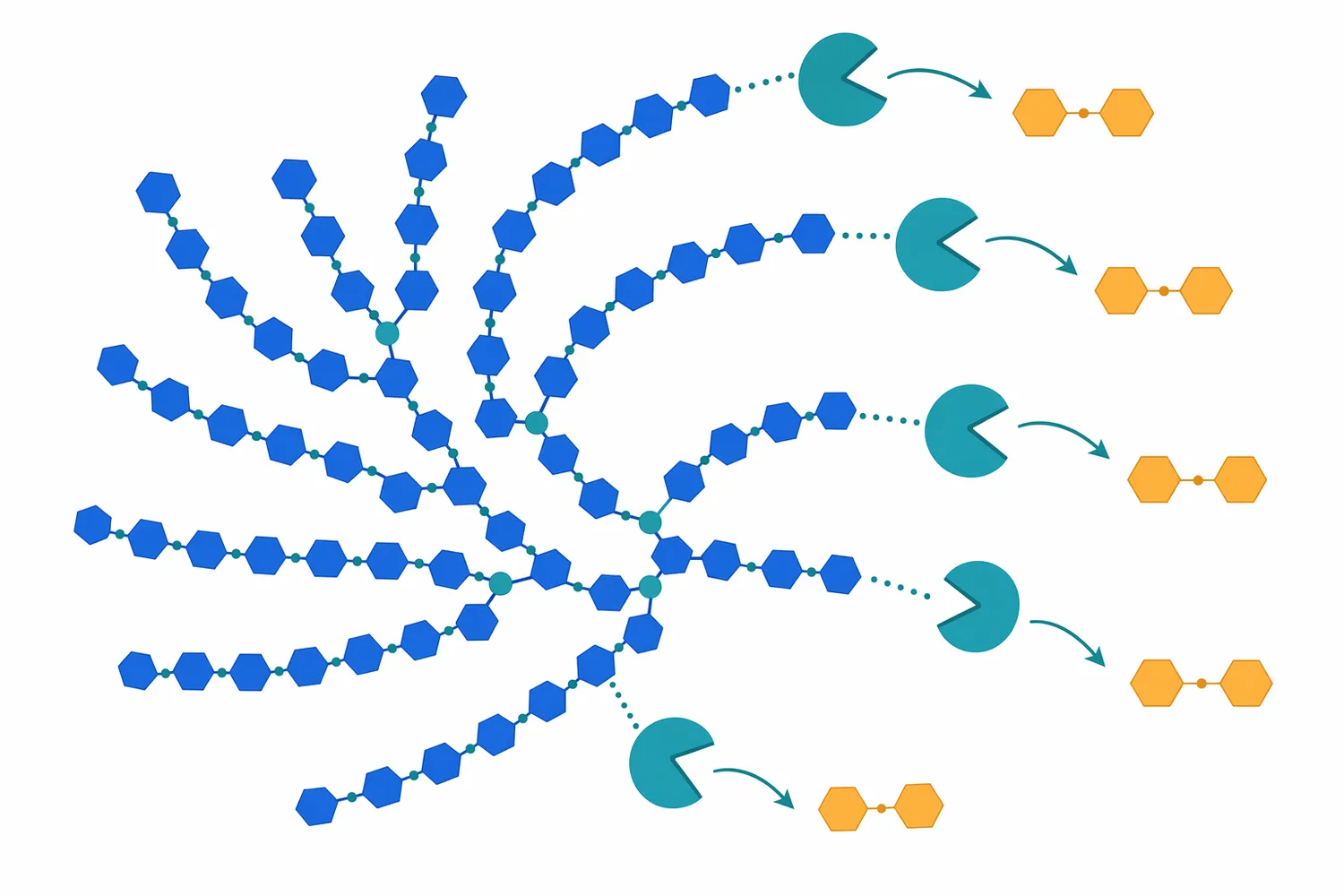
β-amylase를 정확히 쓰려면 “무엇을 잘하는가”뿐 아니라 “어디서 멈추는가”를 알아야 합니다. β-amylase는 α-1,4 결합을 따라 maltose를 방출하지만, α-1,6 가지 결합을 만나면 진행이 제한됩니다. 이때 남는 β-limit dextrin은 불완전 분해의 부산물로 보일 수도 있지만, 전분 구조를 해석하는 분석적 지표로도 사용됩니다 [2].
고 maltose 목표가 높을수록 debranching enzyme의 의미가 커집니다. pullulanase나 limit dextrinase는 amylopectin의 가지 결합 주변 구조를 풀어 β-amylase가 더 긴 α-1,4 구간을 따라 진행할 수 있게 해줍니다. mashing 모델에서 limit dextrinase가 α-amylase 및 β-amylase와 함께 전분 분해에 기여하는 것으로 다루어진 이유도 여기에 있습니다 [6].
실제 공정에서는 “β-amylase 단독”과 “α-amylase + β-amylase”, “β-amylase + pullulanase”, “α-amylase + β-amylase + debranching enzyme”의 결과가 서로 다를 수 있습니다. 조합을 늘린다고 항상 경제적·품질적으로 유리한 것은 아니지만, maltose 비율과 잔류 dextrin 비율을 동시에 관리해야 하는 경우에는 효소 간 기능 분업이 핵심 설계 요소가 됩니다 [9].
β-amylase는 maltose 생산으로 가장 잘 알려져 있지만, 전분 전환 네트워크 안에서는 다른 당류 생산 공정과 연결되기도 합니다. 예를 들어 thermostable beta-amylase와 trehalose synthase 기능을 결합한 bifunctional enzyme 연구는 전분에서 trehalose로 이어지는 전환을 쉽게 만들기 위한 접근으로 보고되었습니다. 이는 β-amylase가 직접 최종 제품을 만드는 효소일 수도 있고, 후속 효소 반응을 위한 maltose성 중간체를 공급하는 역할일 수도 있음을 보여줍니다 [10].
또한 전분을 열적·효소적으로 처리해 resistant maltodextrin을 만드는 연구에서는 전분 구조 변화와 효소 가수분해가 소화 저항성, 물성, 영양 특성에 영향을 줄 수 있음이 보고되었습니다. cassava starch의 pyroconversion 및 enzymatic hydrolysis 연구는 전분 가공에서 “효소 처리”가 단순 당화뿐 아니라 저항성 탄수화물 소재 설계와도 연결될 수 있음을 보여줍니다 [11].
다만 resistant maltodextrin이나 기능성 탄수화물 소재의 경우, β-amylase가 항상 중심 효소라고 단정할 수는 없습니다. 열변성, 산 처리, 다른 amylase, transglycosylation, 분획 조건이 함께 작용하기 때문입니다. 따라서 beta-amylase는 전분 기반 당류·올리고당 공정의 한 축으로 이해하되, 최종 소재 기능성은 전체 공정 이력에 의해 결정된다고 보는 것이 안전합니다 [12].
“beta amylase molecular weight”는 기술 자료를 읽을 때 자주 검색되는 표현입니다. 그러나 β-amylase의 분자량은 단일한 보편값으로 다루기보다, 효소 원천과 단백질 형태에 따라 달라질 수 있는 특성으로 보는 것이 적절합니다. 식물 유래, 미생물 유래, 재조합 효소, 융합 효소는 아미노산 서열과 구조적 도메인 구성이 다를 수 있으며, 이런 차이는 안정성·기질 결합·공정 적합성에 영향을 줄 수 있습니다 [4].
특히 재조합 또는 bifunctional enzyme에서는 β-amylase 기능 도메인에 다른 효소 기능이 연결될 수 있습니다. 전분에서 trehalose 전환을 촉진하기 위해 thermostable beta-amylase-trehalose synthase bifunctional enzyme을 구성한 연구는, “β-amylase”라는 기능명이 항상 동일한 단백질 크기나 동일한 구조를 의미하지 않는다는 점을 잘 보여줍니다 [10].
따라서 기술 문서에서 분자량 정보가 등장하면, 그 값은 특정 효소 원천과 특정 제형 또는 특정 연구 단백질에 대한 정보로 읽어야 합니다. 공정 설계에서 더 직접적으로 중요한 것은 maltose 생성 기능, pH·온도 안정성, 기질 접근성, 다른 효소와의 상호작용이며, 분자량은 그 기능을 해석하는 보조 정보에 가깝습니다 [4].
전분 원료가 바뀌면 beta-amylase activity의 실질적 결과도 달라질 수 있습니다. 옥수수 전분, waxy corn starch, cassava starch, 수수 전분, 보리·맥아 전분은 amylose/amylopectin 구성, granule 크기, gelatinization 특성, 단백질·지질 복합체 여부가 다릅니다. waxy corn starch에서 β-limit dextrin을 준비한 연구는 amylopectin이 많은 원료에서 가지 구조가 β-amylase 반응의 잔류 구조를 만든다는 점을 잘 보여줍니다 [2].
카사바 전분은 resistant maltodextrin 연구에서 자주 다루어지는 원료 중 하나입니다. pyroconversion과 enzymatic hydrolysis가 indigestible starch content와 물리화학적 특성에 영향을 준다는 연구는, 같은 전분이라도 열적 전처리와 효소 처리 조합에 따라 소화성·점도·분자 구조가 달라질 수 있음을 보여줍니다 [11].
수수와 같은 곡물 원료는 발효 공정과 연결될 때 원료의 전분 접근성뿐 아니라 단백질 매트릭스, 입자 분쇄도, 당화액의 영양 조성까지 고려해야 합니다. 수수 곡물에서 에탄올 생산을 평가한 연구는 효소 가수분해와 hydrolysate fermentability를 함께 다루며, 전분을 당으로 바꾸는 단계와 그 당을 실제로 발효하는 단계가 서로 연결되어 있지만 동일하지는 않다는 점을 시사합니다 [8].
β-amylase를 다룰 때 pH, 온도, 반응 시간은 당연히 중요합니다. 그러나 더 먼저 정의해야 할 것은 목표 당 조성입니다. maltose를 최대로 높이고 싶은지, 잔류 dextrin을 일부 유지해 점도와 바디감을 남기고 싶은지, 효모 발효를 위해 glucose와 maltose 균형을 맞추고 싶은지에 따라 같은 beta-amylase라도 적용 의미가 달라집니다 [3].
“beta amylase temperature range”를 단독으로 찾는 접근은 실무적으로 한계가 있습니다. β-amylase가 작용하기 쉬운 온도라도 전분이 충분히 gelatinization되지 않았다면 기질 접근성이 낮을 수 있고, 반대로 전분은 잘 풀렸지만 효소가 빠르게 열실활되는 조건이라면 maltose 생성이 제한될 수 있습니다. mashing의 kinetic model은 효소 반응과 온도 이력이 동시에 전분 분해 결과를 좌우한다는 점을 설명합니다 [1].
pH 역시 효소 안정성과 기질 상태를 함께 바꿉니다. 전분 당화 공정에서는 산성·중성 영역의 미세한 차이가 효소 활성뿐 아니라 전분 분산성, 단백질 침전, 미생물 관리, 후속 발효에 영향을 줄 수 있습니다. maltose syrup 생산 조건을 다룬 연구가 reaction parameter와 process control을 함께 검토한 이유는, 최종 제품 품질이 단일 변수보다 공정 변수의 조합에 의해 결정되기 때문입니다 [3].
β-amylase의 실질적 가치는 전분을 단순히 작게 자르는 데 있지 않습니다. 전분을 어떤 당으로, 어느 정도의 dextrin 잔류를 남기며, 어떤 발효성 또는 감미 특성으로 전환할지 설계하는 데 있습니다. 이 관점에서 beta-amylase function은 공정의 “당화 방향성”을 정하는 효소 기능으로 이해하는 것이 좋습니다 [6].
식품 산업에서 효소는 대개 최종 제품의 물성, 풍미, 영양, 공정성에 영향을 주는 보조 가공 도구로 사용됩니다. microbial enzymes와 food industry 응용을 다룬 최근 리뷰에서도 효소는 반응 특이성을 통해 기존 열·화학 공정보다 선택적인 변환을 제공하는 기술로 정리됩니다. β-amylase도 이 틀 안에서 maltose 중심 전분 전환을 가능하게 하는 선택적 도구로 볼 수 있습니다 [13].
다만 효소 사용 결과를 과장해서는 안 됩니다. β-amylase는 전분 원료의 구조적 한계를 없애지 못하며, α-1,6 가지 결합을 해결하려면 debranching enzyme의 도움이 필요할 수 있습니다. 또한 발효 수율, 시럽 감미, 최종 점도는 β-amylase activity뿐 아니라 전분 전처리, 효소 조합, 반응 종료 시점, 후속 공정에 의해 함께 결정됩니다 [9].
Enzymes.bio는 Beta-Amylase를 제조하는 실험실이나 제조사가 아니라, 효소 원료를 온라인으로 공급하는 B2B 공급업체입니다. 본 제품은 1kg 단위로 온라인에서 직접 구매할 수 있으며, 주문 시 CoA와 SDS가 함께 제공됩니다. 이 문서는 제조 지침이나 성능 보증서가 아니라, beta-amylase의 기전과 응용을 이해하기 위한 기술 설명 자료입니다.
구매자가 이 효소를 검토할 때 핵심은 “전분을 얼마나 분해할 것인가”보다 “maltose 중심의 당 조성이 필요한가”입니다. 고 maltose 시럽, 맥아·곡물 mashing, 발효 가능한 당 조성 설계, 전분 구조 연구처럼 maltose 방출과 β-limit dextrin 형성이 의미 있는 공정이라면 β-amylase가 적합한 후보가 될 수 있습니다 [3].
반대로 전분 액화, 급격한 점도 저감, glucose 중심 당화, branch point 제거가 주목표라면 α-amylase, glucoamylase, pullulanase, limit dextrinase 등 다른 효소와의 역할 구분이 필요합니다. “alpha amylase beta amylase”를 함께 검토하는 이유는 두 효소가 경쟁 관계라서가 아니라, 전분 사슬 내부 절단과 maltose 방출이라는 서로 다른 기능을 맡기 때문입니다 [6].
Beta-Amylase는 α-글루칸의 비환원 말단에서 maltose를 순차적으로 방출하는 exo형 amylase입니다. 이 때문에 고 maltose 시럽, 맥아 당화, 곡물 발효, 전분 구조 연구에서 중요한 효소이며, “beta-amylase activity”의 실무적 의미는 maltose 생성 방향으로 당 조성을 조절하는 능력입니다 [1].
α-amylase와의 차이는 명확합니다. α-amylase는 전분 내부 절단과 액화에 강하고, β-amylase는 말단 절단과 maltose 축적에 강합니다. 두 효소를 조합하면 α-amylase가 만든 새로운 사슬 말단을 β-amylase가 이용할 수 있으며, limit dextrinase나 pullulanase를 더하면 amylopectin 가지 구조로 인한 제한을 완화할 수 있습니다 [9].
β-amylase의 한계도 분명합니다. α-1,6 branch point를 직접 광범위하게 제거하지 못하고, 전분 granule 접근성이 낮으면 반응이 제한되며, 발효 성능은 maltose 형성만으로 결정되지 않습니다. 따라서 beta-amylase는 만능 전분 분해제가 아니라, maltose 중심 전분 전환을 설계하는 특이적 효소로 이해하는 것이 가장 정확합니다 [2].
1kg 단위로 판매되며 재고 보유, 즉시 출고됩니다. 온라인 스토어에서 바로 결제하시면 주문을 처리해 드립니다. 모든 주문에는 시험성적서(CoA)와 물질안전보건자료(SDS)가 포함됩니다.
Beta-Amylase 구매하기 →최초 인용 순서로 번호를 매겼습니다. 모든 출처는 발행 시점에 접근 가능 여부를 확인한 오픈 액세스 자료이며, 본문의 인용 번호가 이곳으로 연결됩니다.