Beta-Glucosidase(β-グルコシダーゼ)は、β-D-グルコシド結合を加水分解し、セロビオースや各種β-グルコシドからグルコースを放出する酵素です。セルロース系バイオマスの糖化では、セルラーゼ反応で蓄積しやすいセロビオースをグルコースへ変換することで、糖化の最終段階を支えます[1]。また、植物配糖体、発酵素材、食品・飲料原料の処理では、糖が結合した成分をアグリコン側へ変換する用途にも利用されます[2]。
Enzymes.bioは製造業者または研究所ではなく、業務用酵素をオンラインで供給するサプライヤーです。Beta-Glucosidaseは1 kg単位でオンラインから直接購入でき、注文時にはCoAおよびSDSが併せて提供されます。
Beta-Glucosidaseは、β-D-グルコース残基を含むグリコシド結合を加水分解する酵素で、セルロース分解系では一般にcellobiaseとしても扱われます。セルロース分解は、結晶性または非晶性セルロースを短鎖オリゴ糖へ崩す段階と、それらを発酵性糖へ仕上げる段階に分けて理解できますが、β-グルコシダーゼは主に後者、すなわちセロビオースや可溶性セロオリゴ糖をグルコースへ変換する末端工程で重要です[3]。
この酵素の実務上の価値は、「大きな多糖を直接壊す主役」というより、セルロース分解で生じる中間生成物を取り除き、反応全体を止まりにくくする点にあります。セロビオースが残ると、セルロースに作用する他のセルラーゼの反応が抑制されやすくなるため、β-グルコシダーゼの不足はリグノセルロース糖化のボトルネックとして繰り返し指摘されています[1]。
β-グルコシダーゼは微生物、真菌、植物など広い生物群に存在し、産業用途では真菌由来および微生物由来の酵素が多く研究されています。特にリグノセルロース資源の利用では、セルラーゼ、ヘミセルラーゼ、補助酵素と組み合わせた糖化系の中で、β-グルコシダーゼの配合や安定性が加水分解効率に大きく影響します[4]。
セルロース分解では、エンドグルカナーゼがセルロース鎖の内部に切れ目を入れ、セロビオヒドロラーゼが鎖端からセロビオースを放出し、β-グルコシダーゼがそのセロビオースをグルコースへ加水分解します。この三段階は単なる直列反応ではなく、各段階の生成物が次の段階の基質になり、かつ蓄積物が前段階を阻害するため、酵素間のバランスが反応速度に直結します[5]。
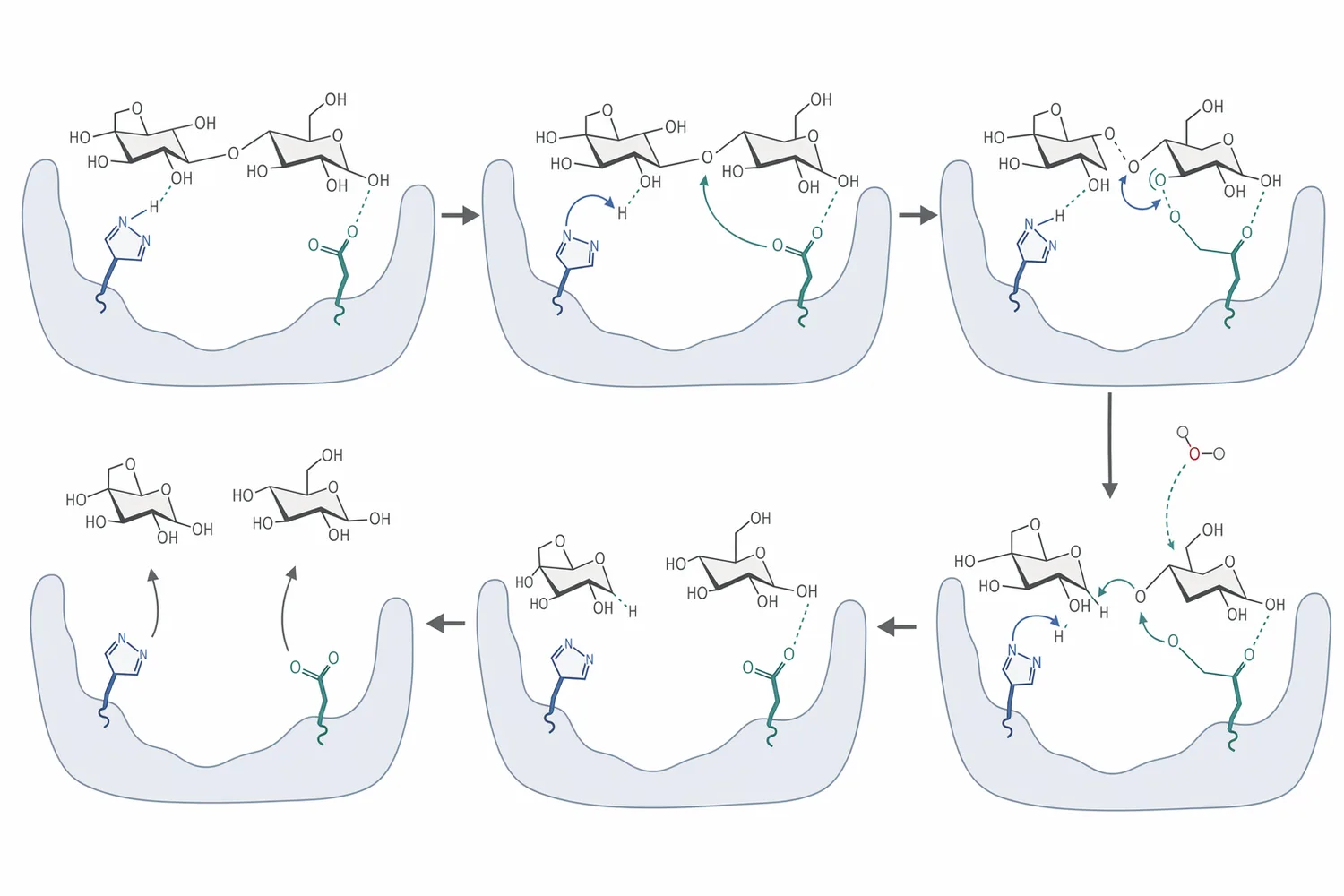
β-グルコシダーゼが十分に機能すると、セロビオース濃度が下がり、セロビオヒドロラーゼやエンドグルカナーゼが生成物阻害を受けにくくなります。一方で、β-グルコシダーゼが不足すると、セルロース表面で生成したセロビオースが反応液中に蓄積し、セルラーゼ複合系の見かけの活性が低下します。このため、β-グルコシダーゼはセルロース加水分解における「仕上げ酵素」であると同時に、上流のセルラーゼ反応を間接的に支える制御点でもあります[6]。
分子レベルでは、多くのβ-グルコシダーゼは糖質加水分解酵素ファミリーに属し、活性部位で基質のβ-グルコシド結合を配置し、酸塩基触媒と求核触媒によりグリコシド結合を切断します。反応は、基質認識、グリコシド結合の活性化、水分子による加水分解、グルコース放出という流れで進みますが、基質の立体構造、糖鎖長、アグリコン部分の疎水性、反応液中の生成物濃度によって速度が変わります[3]。
リグノセルロース系原料には、セルロース、ヘミセルロース、リグニンが複雑に絡み合っています。セルロース鎖そのものはグルコースのβ-1,4結合ポリマーですが、天然の植物細胞壁ではリグニンとヘミセルロースが酵素のアクセスを制限するため、セルラーゼ系酵素だけでなく前処理、補助酵素、界面現象まで含めた設計が必要になります[5]。
β-グルコシダーゼは、前処理後に可溶化または露出したセルロース鎖から生じるセロビオースを処理する役割を担います。特に高固形分の糖化では、基質濃度が高いほど生成物も蓄積しやすく、グルコースやセロビオースによる生成物阻害が収率を左右する要因になります。トウモロコシストーバーの高固形分加水分解に関する研究でも、生成物阻害が糖化収率を決める重要因子として扱われています[6]。
セルロース糖化におけるβ-グルコシダーゼの効果は、単独の酵素活性だけでは評価しきれません。セルロース表面への吸着、リグニンへの非生産的吸着、他のセルラーゼとの比率、反応液中の糖濃度が重なって、最終的なグルコース生成量が決まります。したがって、β-グルコシダーゼは「投入すれば常に同じ効果が出る添加物」ではなく、セルラーゼ系全体の律速点を補う構成要素として理解する必要があります[1]。
| 酵素区分 | 主な作用点 | 主な生成物・効果 | β-グルコシダーゼとの関係 |
|---|---|---|---|
| エンドグルカナーゼ | セルロース鎖内部 | 新しい鎖端、短鎖セルロオリゴ糖 | 鎖を開裂して後続反応の入口を作る |
| セロビオヒドロラーゼ | セルロース鎖端 | セロビオース | β-グルコシダーゼの主要基質を供給する |
| β-グルコシダーゼ | セロビオース、短鎖β-グルコシド | グルコース | セロビオースを除去し、生成物阻害を緩和する |
| 補助酵素・関連酵素 | ヘミセルロース、リグニン周辺結合など | 基質露出、構造緩和 | β-グルコシダーゼが働く可溶性糖の供給を助ける |
この比較から分かるように、β-グルコシダーゼはセルロース鎖を最初に開く酵素ではありません。しかし、最終糖化で詰まりやすいセロビオースを処理するため、少量の不足でも全体の糖化速度が下がることがあります。レビューでは、真菌β-グルコシダーゼの性能と供給がリグノセルロース利用の産業化における重要課題として位置づけられています[3]。
リグノセルロース糖化でβ-グルコシダーゼを考える場合、基質は精製セルロースだけではありません。実際のバイオマスにはリグニン由来成分が残り、これが酵素を非生産的に吸着したり、セルラーゼ、キシラナーゼ、β-グルコシダーゼの活性を阻害したりします。軟材リグニン調製物を用いた研究では、セルラーゼ、キシラナーゼ、β-グルコシダーゼ活性に対するリグニンの阻害が検討されており、木質系原料ではリグニンの影響を無視できないことが示されています[7]。
リグニンの影響は一方向ではなく、リグニンの化学構造、スルホン化の程度、前処理条件、酵素表面の性質によって変わります。リグノスルホン酸ナトリウムがセルロース酵素糖化に与える影響を調べた研究では、基質と酵素の吸着挙動の観点から、添加物が酵素の非生産的吸着を変化させ得ることが議論されています[8]。
軟材のように難分解性の高い原料では、前処理がβ-グルコシダーゼの働きやすさを大きく左右します。SPORLと呼ばれる亜硫酸前処理をトウヒおよびレッドパインに適用した研究では、堅牢な酵素糖化を目指して、リグニンの改変と繊維構造の変化が糖化性に与える影響が検討されています[9]。
近年は、リグニンやヘミセルロース周辺の結合を変える補助酵素にも関心が向けられています。グルクロノイルエステラーゼがリグノセルロース分解酵素によるセルロース加水分解を改善し、リグニン抽出を高めることを示した研究は、β-グルコシダーゼ単独ではなく、細胞壁構造全体をほどく酵素ネットワークの重要性を示しています[10]。

β-グルコシダーゼは、バイオエタノールやバイオベース化学品の原料となる発酵性糖を作る工程で重要です。セルロースを含む農業残渣、木質原料、都市系紙資源などは、前処理と酵素糖化を経てグルコースなどの糖に変換され、その後の発酵工程に供給されます。微生物β-グルコシダーゼに関するレビューでは、バイオ燃料用途でのバイオマス変換における同酵素の役割が整理されています[4]。
廃紙や都市固形廃棄物由来の紙資源は、セルロースを多く含むため、糖化・発酵の対象として研究されてきました。都市固形廃棄物および関連する廃紙流をバイオアルコール生産に活用する研究では、前処理、酵素糖化、発酵を組み合わせることで、廃棄物由来炭素を液体燃料へ転換する考え方が示されています[11]。
ただし、β-グルコシダーゼの追加が常に単独で収率を決めるわけではありません。原料がリグニンを多く含む場合、酵素が基質に届かない、または非生産的に吸着することがあります。高固形分条件では、糖濃度上昇による生成物阻害も強くなります。したがって、β-グルコシダーゼはセルラーゼ配合、前処理、固形分、糖化時間、後段発酵との接続を含めた工程設計の中で意味を持ちます[6]。
植物原料には、香気前駆体、フェノール性化合物、イソフラボン、サポニン様成分など、糖が結合した配糖体が多く存在します。β-グルコシダーゼはこれらのβ-グルコシド結合を切断し、グルコースとアグリコンを放出するため、植物抽出物、発酵食品、飲料原料の処理に利用されます。ワイン酵母のβ-グルコシダーゼ活性に関するレビューでは、ワイン中の揮発性成分やフェノール性成分への影響が議論されています[2]。
ワインや果実発酵では、テルペン系香気成分などが糖結合型で存在することがあり、β-グルコシダーゼ反応によって香気前駆体から揮発性アグリコンが遊離する可能性があります。ただし、香気への影響は基質、酵母株、pH、エタノール、糖、有機酸、発酵温度に左右されるため、酵素反応だけから官能品質を一律に保証することはできません[2]。
大豆イソフラボンでは、配糖体型からアグリコン型への変換が食品加工上の関心対象になります。β-グルコシダーゼの固定化と大豆イソフラボン配糖体の加水分解を扱った研究では、酵素を用いた配糖体変換が検討されており、植物性機能成分の加工にβ-グルコシダーゼが関与し得ることを示しています[12]。
発酵植物素材では、微生物が産生するリグノセルロース分解酵素の組み合わせにより、細胞壁分解と配糖体変換が同時に進むことがあります。Auricularia auriculaを用いたトウモロコシ茎残渣の固体発酵研究では、前処理条件によって酵素産生とリグノセルロース分解パターンが変わることが示され、β-グルコシダーゼを含む酵素群の働きが発酵原料の変化に関係することが分かります[13]。
β-グルコシダーゼは水溶性酵素として利用されることが多い一方で、研究開発では固定化による再利用性、安定性、反応制御の改善も検討されています。Aspergillus versicolor由来β-グルコシダーゼの生産、固定化、特性評価を扱った研究では、セルロース分解への応用を視野に入れ、固定化による酵素利用の可能性が検討されています[14]。
セルラーゼとβ-グルコシダーゼを同じ担体に共固定化する研究もあります。メソポーラスシリカナノ粒子にセルラーゼとβ-グルコシダーゼを共固定化し、ビワ葉由来セルロースの加水分解に適用した研究では、複数酵素を近接配置することで、セルロース分解からセロビオース加水分解までの連続反応を効率化する考え方が示されています[15]。
固定化の利点は、酵素の回収や連続反応への組み込みだけではありません。担体表面の親水性、細孔径、酵素の向き、拡散距離が反応性に影響するため、酵素を「溶液中に入れる」場合とは異なる速度論が生じます。一方で、基質が大きい場合は担体内部へ入りにくく、拡散制限が反応を遅くすることもあるため、固定化は用途に応じた選択肢として扱うべきです[15]。
Enzymes.bioの供給するBeta-Glucosidaseは、オンラインで購入できる業務用酵素として位置づけられます。固定化、共固定化、複合酵素系への組み込みは、各社の工程設計や研究開発のテーマであり、供給品そのものを製造プロセスや研究成果と同一視するものではありません。
β-グルコシダーゼの実用性を左右する主要因の一つが生成物阻害です。多くのβ-グルコシダーゼでは、生成物であるグルコースが活性部位または関連結合部位に影響し、セロビオース加水分解を抑えることがあります。Aspergillus niger由来の商用β-D-グルコシダーゼ調製物を対象にした古典的研究では、速度論、阻害、安定性の観点から、リグノセルロース加水分解への適性が検討されています[16]。
一方で、すべてのβ-グルコシダーゼが同じようにグルコースで阻害されるわけではありません。近年は、グルコース存在下でも働きやすい、あるいはグルコースによって活性が刺激されるβ-グルコシダーゼの改良研究も進んでいます。Bgl2Aのセロビオース加水分解活性を改善した研究では、グルコース刺激型β-グルコシダーゼの性質を活かし、生成物存在下での反応性向上が検討されています[17]。
ただし、グルコース耐性だけで糖化性能が決まるわけではありません。リグニンへの非生産的吸着、前処理由来阻害物質、pH、温度、イオン強度、基質濃度、他酵素との相互作用が重なります。リグニン調製物によるβ-グルコシダーゼ活性阻害の研究は、精製基質で見える酵素性能と、実バイオマスで得られる糖化性能に差が出る理由を説明する根拠になります[7]。
リグノセルロース糖化では、酵素と基質の接触を改善するために、界面活性剤、バイオサーファクタント、有機アルコールなどの添加効果が研究されています。これらは酵素の直接活性を上げるというより、リグニンへの非生産的吸着を減らしたり、基質表面の濡れ性や酵素分布を変えたりすることで、加水分解効率に影響する場合があります[18]。
有機アルコールとバイオサーファクタントの複合添加によるリグノセルロース酵素加水分解の相乗的向上を扱った研究では、添加物の組み合わせが糖化効率に影響し得ることが示されています。β-グルコシダーゼにとっても、セロビオースがどれだけ供給され、生成物がどれだけ蓄積し、酵素がどれだけ反応液中に自由に存在できるかが重要です[18]。
ただし、食品、発酵、バイオマス、飼料、化学原料などの用途では、許容される添加物や工程条件が異なります。したがって、β-グルコシダーゼの利用は、最終用途の規格、工程の温度域、反応後の処理、他成分との適合性を含めて設計される必要があります。
β-グルコシダーゼの研究では、発現量、熱安定性、pH安定性、グルコース耐性、基質特異性の改善を目的とした酵素工学が進められています。バイオ燃料用途に向けたβ-グルコシダーゼ生産強化に関するレビューでは、遺伝子改変が酵素生産性や特性改善の手段として整理されています[19]。
酵素工学の目的は、単に「活性を高くする」ことだけではありません。高固形分糖化では生成物濃度が高くなるため、グルコース存在下で働けることが重要になります。木質原料や農業残渣では前処理由来化合物が残るため、阻害物質への耐性も関係します。さらに、実工程では糖化時間が長くなることがあり、反応中の安定性も重要です[4]。
一方で、研究論文で示された特定酵素の改良結果を、すべての市販β-グルコシダーゼに一般化することはできません。由来生物、配列、糖鎖修飾、精製度、配合、担体、保存形態が異なれば、同じ「β-グルコシダーゼ」という名称でも挙動は変わります。したがって、酵素工学の知見は、β-グルコシダーゼ利用の方向性を理解する根拠として扱うのが適切です[19]。
| 用途領域 | 主な基質・対象 | β-グルコシダーゼの役割 | 期待される工程上の価値 | 注意点 |
|---|---|---|---|---|
| セルロース糖化 | セロビオース、短鎖セロオリゴ糖 | グルコースへの最終加水分解 | セロビオース蓄積の低減、発酵性糖生成 | リグニン阻害、生成物阻害、他酵素との比率に依存 |
| バイオ燃料 | 農業残渣、木質原料、廃紙系資源 | セルラーゼ系の補完 | 糖化液のグルコース化、発酵原料化 | 前処理条件と高固形分阻害が重要 |
| 食品・飲料 | 植物配糖体、香気前駆体 | グルコースとアグリコンの遊離 | 香気・成分変換の可能性 | 官能結果は原料と工程に依存 |
| 植物抽出物 | イソフラボンなどの配糖体 | アグリコン型への変換 | 抽出物の成分プロファイル調整 | 健康効果を酵素反応だけで主張しない |
| 発酵素材 | 植物細胞壁、可溶性β-グルコシド | 糖供給、配糖体変換 | 微生物利用可能糖の増加 | 発酵微生物と条件の影響が大きい |
| 固定化反応 | 可溶性β-グルコシド、セルロース由来糖 | 反復利用・連続化の検討 | 酵素回収、反応制御 | 拡散制限や担体適合性が課題 |
この表が示すように、Beta-Glucosidaseの価値は「グルコースを作る」だけではありません。セルラーゼ系の阻害緩和、植物配糖体の変換、発酵原料の糖組成調整、固定化プロセスへの応用など、用途ごとに異なる機能が前面に出ます。特にセルロース糖化では、β-グルコシダーゼの不足が全体効率の制限要因になり得るため、セルラーゼ配合の中核的な補完酵素として扱われます[1]。
β-グルコシダーゼ反応は、基質濃度、温度、pH、水分、生成物濃度、共存する糖、有機酸、塩、フェノール性化合物の影響を受けます。食品・飲料では酸性条件や糖濃度、エタノールの存在が問題になり、バイオマス糖化ではリグニン、前処理副生成物、高固形分、粘度が問題になります。したがって、同じ酵素でも、透明なモデル基質での挙動と実原料中での挙動は一致しないことがあります[16]。
反応設計では、β-グルコシダーゼを単独酵素としてではなく、生成物の流れを制御する酵素として捉えると理解しやすくなります。セルロース糖化では、セロビオース生成速度とセロビオース加水分解速度の釣り合いが重要です。植物配糖体変換では、配糖体の溶解性、アグリコンの安定性、共存ポリフェノールとの相互作用が結果に影響します[2]。
また、反応が進むほどグルコース濃度が上がるため、初期速度だけでなく反応後半の失速も考慮する必要があります。高固形分糖化で生成物阻害が収率を左右するという知見は、β-グルコシダーゼを含む酵素系の設計において、糖蓄積をどう扱うかが重要であることを示しています[6]。
Enzymes.bioは、Beta-Glucosidaseをオンラインで供給するサプライヤーです。製品は1 kg単位で直接購入でき、オンライン注文後に処理・配送されます。注文時にはCoAおよびSDSが併せて提供されるため、受領後の社内管理、安全情報確認、品質関連書類の保管に利用できます。
Enzymes.bioは製造業者や研究所ではなく、酵素を業務用途向けに販売する供給事業者です。そのため、本ページで扱う技術情報は、β-グルコシダーゼの用途理解、工程検討、原料処理の考え方を整理するためのものであり、特定原料での最終性能や完成品の効果を保証するものではありません。
Beta-Glucosidaseは、β-D-グルコシド結合を加水分解し、セロビオースや植物配糖体からグルコースを放出する酵素です。セルロース糖化では、セロビオースをグルコースへ変換することで、セルラーゼ系全体の反応を支え、生成物阻害の緩和に関与します[1]。
リグノセルロース用途では、β-グルコシダーゼの効果はリグニン、前処理、基質吸着、生成物阻害、他酵素との組み合わせに左右されます。食品・飲料・植物抽出物では、配糖体からアグリコンを遊離させる反応が価値を持ちますが、香味、機能性、発酵挙動は原料と工程に依存します[2]。
Enzymes.bioのBeta-Glucosidaseは、セルロース系原料処理、発酵素材、植物配糖体変換、食品・飲料原料の加工検討などに利用できる業務用酵素として、1 kg単位でオンライン購入できます。CoAおよびSDSは注文時に併せて提供されます。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Beta-Glucosidaseを購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。