Beta-glucosidase, beta-glikozidik bağ taşıyan glukozitleri su kullanarak glukoza ve bağlı aglikon yapıya ayıran bir enzimdir; endüstriyel açıdan en yerleşik kullanım alanı lignoselülozik biyokütle sakkarifikasyonunda sellobiyoz ve kısa glukozit ara ürünlerinin glukoza dönüştürülmesidir [1]. Aynı bağ kesme mantığı, gıda ve bitkisel içerik proseslerinde bağlı aroma öncülleri, fenolik glikozitler ve bazı özel glukozitlerin dönüştürülmesi için de teknik olarak önemlidir [2]. Enzymes.bio, Beta-Glucosidase ürününü üretici veya laboratuvar olarak değil tedarikçi olarak sunar; ürün çevrim içi olarak 1 kg birimler halinde doğrudan satın alınır ve siparişle birlikte CoA ile SDS sağlanır.
Beta-glucosidase, molekül üzerinde glukoza bağlı beta-glikozidik bağ bulunan substratları hedefleyen bir biyokatalizördür. Reaksiyonun sade karşılığı şudur: Enzim, glukozit yapıyı aktif bölgesinde konumlandırır, beta-glikozidik bağı su yardımıyla kırar ve sonuçta serbest glukoz ile glukoza bağlı olan ikinci parçayı, yani aglikon veya daha küçük karbonhidrat parçasını açığa çıkarır [3].
Bu özellik özellikle sellobiyoz için kritiktir. Selüloz parçalanırken daha büyük zincirler önce kısa selüloz türevli oligosakkaritlere ve sellobiyoza ayrılır; beta-glucosidase bu ara ürünleri glukoza kadar ilerleten son basamakta çalışır. Bu nedenle beta-glucosidase, selülaz sistemiyle birlikte düşünüldüğünde “küçük ara ürünleri temizleyen” tamamlayıcı enzim olarak değerlendirilir [1].
Beta-glucosidase yalnızca selüloz bazlı ara ürünlerle sınırlı değildir. Bitkisel hammaddelerde glukozla bağlı fenolikler, flavonoid glikozitleri veya aromatik aglikonlar bulunabilir; glukozun kopması bu bileşiklerin çözünürlüğünü, reaktivitesini, duyusal etkisini veya biyolojik erişilebilirliğini değiştirebilir. Pelargonidin-3-O-glucoside gibi antosiyanin glikozitlerinin gıda matriksindeki etkileşimleri ve biyolojik erişilebilirliği üzerine çalışmalar, glikozit formunun proses ve formülasyon açısından ayrı bir kimyasal davranış gösterebildiğini ortaya koyar [4].
Bu noktada beta-glucosidase ile beta-glucan veya beta-glucanase terimleri karıştırılmamalıdır. Beta-glucan, yulaf, arpa, maya veya mantar kaynaklı çözünür lif olarak ele alınan polisakkarit bir bileşendir; beta-glucosidase ise belirli glukozit bağlarını kesen enzimdir. Beta-glucan literatürü daha çok çözünür lifin kaynağı, yapısı, ekstraksiyonu, biyoyararlanımı ve gıda uygulamalarıyla ilgilenir [5].
Beta-glucosidase’in pratik mekanizması üç aşamada anlaşılabilir: substratın tanınması, glikozidik bağın zayıflatılması ve suyla hidroliz. Enzim aktif bölgesi, glukoz kısmını belirli bir geometriyle tutar; bu konumlandırma, beta-glikozidik bağın kimyasal olarak kırılmaya uygun hale gelmesini sağlar. Aspergillus oryzae beta-glucosidase üzerinde yapılan bağlanma enerjisi ve mekanizma çalışmaları, substratın aktif bölgede doğru tutulmasının reaksiyonun verimliliği için temel unsur olduğunu göstermiştir [3].
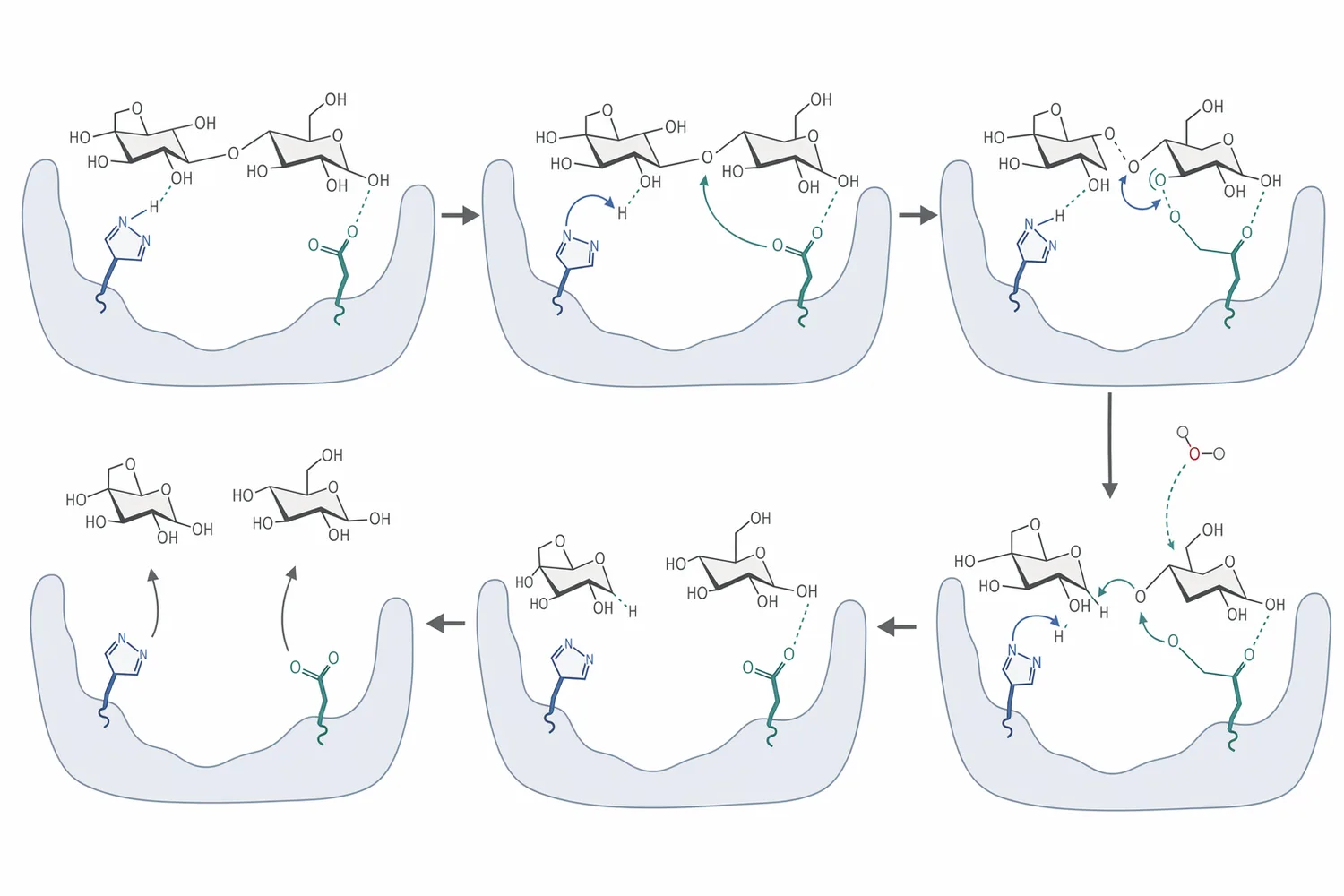
Birçok beta-glucosidase örneğinde reaksiyon, glukozil-enzyme ara durumu üzerinden ilerleyen “tutucu” mekanizma olarak açıklanır: önce glikozidik bağ kırılır ve glukoz kısmı enzimle geçici olarak ilişkilendirilir; ardından su molekülü devreye girerek glukozu serbestleştirir. Thermoascus aurantiacus kaynaklı termostabil beta-glucosidase üzerine yapılan biyokimyasal ve mekanistik çalışma, bu tür enzimlerde aktif bölge kimyasının hidroliz davranışını belirlediğini gösteren klasik örneklerdendir [2].
Aktif merkezin doğrudan incelendiği eski çalışmalar da mekanizmanın somutlaşmasına katkı sağlamıştır. Aspergillus wentii beta-glucosidase aktif merkezinin işaretlenmesi üzerine yapılan çalışma, katalitik bölgenin yalnızca pasif bir bağlanma cebi olmadığını; bağ kesme reaksiyonunda doğrudan görev alan özel bir kimyasal ortam sunduğunu ortaya koymuştur [6].
Bu mekanizma, farklı uygulamalarda aynı temel sonucu üretir fakat ürünün anlamı değişir. Sellobiyozda sonuç iki glukoz birimidir; aromatik glikozitte sonuç serbest aroma aglikonudur; flavonoid glikozitte sonuç aglikon flavonoid ve glukozdur. Bu nedenle beta-glucosidase’in endüstriyel değeri, tek bir üründen ziyade “glukoza bağlı halde bulunan bileşiklerin kontrollü biçimde açığa çıkarılması” kabiliyetine dayanır [7].
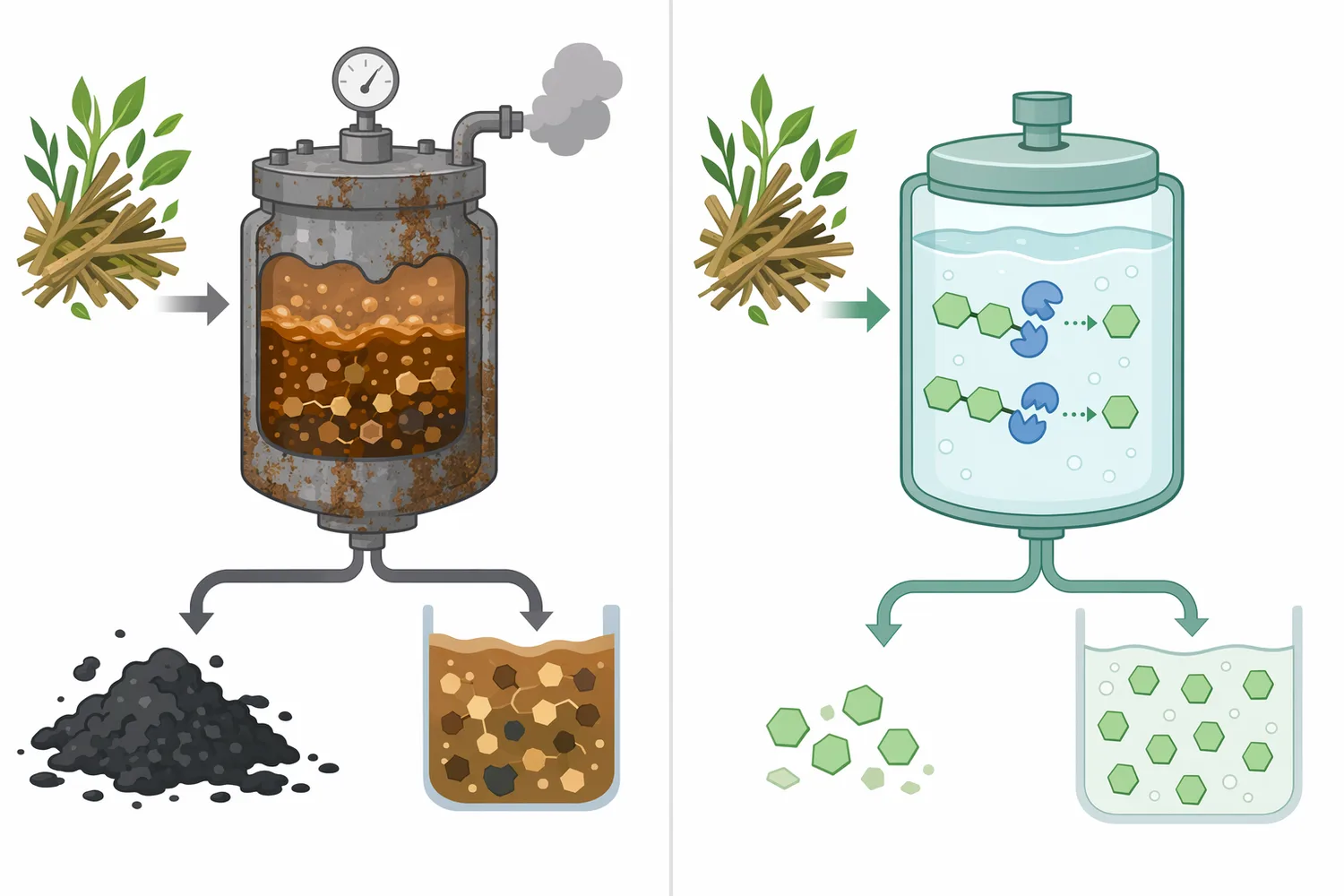
Beta-glucosidase’in en güçlü teknik dayanağa sahip kullanım alanı lignoselülozik biyokütlenin şekere dönüştürülmesidir. Selülozun enzimatik hidrolizinde farklı selülaz bileşenleri selüloz zincirini parçalarken sellobiyoz ve kısa glukozit ara ürünleri oluşur; beta-glucosidase bu ara ürünleri glukoza çevirerek sakkarifikasyonun tamamlanmasına katkı verir [1].
Bu reaksiyon yalnızca ek glukoz üretmek için değil, proses kinetiği açısından da önemlidir. Sellobiyoz birikimi, selülozun daha ileri parçalanmasını sınırlayabilen bir ara ürün olarak ele alınır; beta-glucosidase ara ürünü uzaklaştırarak diğer enzimlerin çalışabileceği kimyasal dengeyi destekler. Aspergillus niger kaynaklı beta-glucosidase’in lignoselülozik biyokütle sakkarifikasyon ürünleri ve fermantasyon inhibitörlerine tolerans bağlamında incelenmesi, bu enzimin endüstriyel biyokütle ortamlarına uyarlanmasının neden önemli olduğunu gösterir [1].
Biyokütle uygulamalarında tek başına beta-glucosidase kullanımı genellikle yeterli bir selüloz dönüşümü anlamına gelmez; çünkü selüloz zincirinin ilk parçalanması için başka enzimler de gerekir. Ancak beta-glucosidase’in eksik veya yetersiz olduğu sistemlerde sellobiyoz ve kısa oligosakkaritlerin birikmesi, toplam şeker verimini ve dönüşüm hızını sınırlayabilir. Bu nedenle beta-glucosidase, selülaz karışımlarının tamamlayıcı ve çoğu zaman kritik bileşeni olarak değerlendirilir [8].
Beta-glucosidase’in fermantasyonla ilişkisi iki yönden önemlidir: fermentasyon öncesi şeker hazırlama ve fermentasyon sırasında glukozitlerin hücre tarafından erişilebilir hale getirilmesi. Endüstriyel diploid maya suşlarında birden fazla beta-glucosidase geninin kromozomal entegrasyonuyla ilgili çalışma, glukozit dönüşüm kapasitesinin canlı hücre bazlı proseslerde de tasarım hedefi olabildiğini göstermektedir [9].
Bu yaklaşım özellikle lignoselülozik hammaddeden yakıt, organik asit veya başka biyobazlı ürün üretiminde anlam kazanır. Enzimatik sakkarifikasyonla oluşan glukoz, uygun mikroorganizma tarafından fermente edilebilir; beta-glucosidase burada sellobiyoz gibi ara ürünlerin glukoza kadar ilerlemesini sağlar. Ancak canlı hücreye entegre beta-glucosidase aktivitesi ile harici enzim kullanımı aynı proses davranışını vermeyebilir; hücre zarı, substrat erişimi, inhibisyon ve ortam koşulları sonucu etkiler [9].
Metagenomik çalışmalarda rumen gibi doğal lignoselüloz parçalayıcı ekosistemlerden beta-glucosidase genlerinin araştırılması da bu nedenle önemlidir. Keçi rumen bakterilerinin metagenom verilerinden beta-glucosidase kodlayan genlerin seçilmesi ve ifade edilmesi üzerine çalışmalar, doğada biyokütle parçalanmasına uyum sağlamış enzim çeşitliliğinin endüstriyel adaylar için kaynak olabileceğini gösterir [10].
Gıda ve içecek proseslerinde beta-glucosidase’in değeri, glukozla bağlı halde bulunan bileşiklerin serbest hale getirilmesinden gelir. Bu bileşikler aromatik aglikonlar, fenolikler, bazı renk bileşikleri veya hammaddenin karakterini etkileyen bitkisel metabolitler olabilir. Glukozit formu çoğu zaman daha farklı çözünürlük, stabilite ve matriks etkileşimi gösterdiği için glukozun kopması nihai ürün profilini değiştirebilir [4].

Pancar, deniz pancarı ve kırmızı pancar literatürü, bitkisel hammaddelerin yalnızca temel besin öğeleriyle değil; betalainler, fenolikler ve farklı biyoaktif bileşenlerle de proses değeri taşıdığını vurgular. Bu tür hammaddelerde enzimatik yaklaşım, renk, tat, fenolik profil veya ekstrakt davranışı gibi değişkenleri etkileyebilen glikozit dönüşümlerinin daha kontrollü yönetilmesine olanak sağlayabilir [11].
Bununla birlikte gıda uygulamalarında beta-glucosidase her matris için aynı sonucu vermez. pH, şeker yoğunluğu, fenolik bileşikler, alkol, tuz, sıcaklık ve hammadde parçacık yapısı enzimin substrata erişimini ve stabilitesini etkileyebilir. Bu nedenle gıda proseslerinde beta-glucosidase, “standart aroma artırıcı” gibi değil; hedef glukozit yapısı ve matris koşulları bilinen bir biyokatalitik araç olarak değerlendirilmelidir [12].
Flavonoidler ve fenolik bileşikler bitkisel hammaddelerde farklı glikozit formlarında bulunabilir. Glikozit formu suda çözünürlüğü ve depolanma stabilitesini etkileyebilir; aglikon formu ise farklı çözünürlük, hücresel etkileşim veya biyolojik erişilebilirlik gösterebilir. Pelargonidin-3-O-glucoside üzerine yapılan biyolojik erişilebilirlik çalışması, tek bir glikozit bileşiğin bile proteinler ve polisakkaritlerle etkileşime girdiğinde farklı davranabildiğini gösterir [4].
Beta-glucosidase bu alanda, glukozit formundan aglikon forma geçişi sağlayabilen araçlardan biridir. Ancak “aglikon her zaman daha iyi ürün demektir” gibi bir genelleme teknik olarak doğru değildir; bazı ürünlerde renk stabilitesi, oksidasyon hassasiyeti veya tat eşiği gibi parametreler aglikon oluşumuyla birlikte değişebilir. Bu nedenle flavonoid dönüşümlerinde hedef, yalnızca glukoz koparmak değil, nihai ürün fonksiyonunu ve raf ömrünü birlikte yönetmektir [13].
Kırmızı pancar ve yan ürünleri üzerine yapılan kapsamlı değerlendirmeler, ekstraksiyon yöntemi, fitokimyasal profil, sağlıkla ilişkilendirilen bileşenler ve uygulama alanları arasında güçlü bir bağlantı olduğunu gösterir. Beta-glucosidase gibi enzimlerin bu tür matrikslerdeki rolü, seçilmiş glikozitlerin dönüştürülmesiyle sınırlı düşünülmeli; hammaddenin tüm fitokimyasal kompozisyonunu tek başına belirleyen bir faktör gibi sunulmamalıdır [13].
| Uygulama alanı | Tipik hedef substrat | Beklenen dönüşüm | Teknik fayda | Dikkat edilmesi gereken sınır |
|---|---|---|---|---|
| Lignoselülozik biyokütle | Sellobiyoz ve kısa selüloz türevli glukozitler | Glukoz oluşumu | Sakkarifikasyonun tamamlanmasına ve ara ürün birikiminin azalmasına katkı | Diğer selülaz bileşenleriyle birlikte düşünülmelidir [1] |
| Fermantasyon öncesi şeker hazırlama | Biyokütle hidroliz ara ürünleri | Fermente edilebilir glukoz | Fermantasyon hammaddesinin daha erişilebilir hale gelmesi | İnhibitörler, glukoz birikimi ve matris etkileri sonucu değiştirebilir [1] |
| Bitkisel ekstrakt ve gıda prosesi | Fenolik veya aroma ilişkili glikozitler | Aglikon + glukoz | Duyusal profil veya bileşen erişilebilirliğinde değişim | pH, fenolikler, alkol ve şeker yoğunluğu aktiviteyi etkileyebilir [12] |
| Flavonoid dönüşümü | Antosiyanin veya flavonoid glikozitleri | Aglikon oluşumu veya glikozit profilinde kayma | Biyolojik erişilebilirlik ve formülasyon davranışı değişebilir | Stabilite, renk ve oksidasyon etkileri ayrıca yönetilmelidir [4] |
| Biyokatalitik sentez | Glukoz donörleri ve uygun alıcı moleküller | Transglikozilasyon ürünleri | Özel glikozit veya oligosakkarit üretim potansiyeli | Su aktivitesi ve substrat dengesi hidrolizle yarışır [7] |
Beta-glucosidase performansı, enzimin kaynağına ve proses ortamına bağlıdır. Fungal, bakteriyel, bitkisel veya rekombinant kaynaklı beta-glucosidase örnekleri farklı sıcaklık, pH, çözücü, iyon ve inhibitör davranışları gösterebilir; bu nedenle literatürde “tek bir beta-glucosidase davranışı” yerine belirli kaynak ve belirli proses koşulları üzerinden yorum yapılır [14].
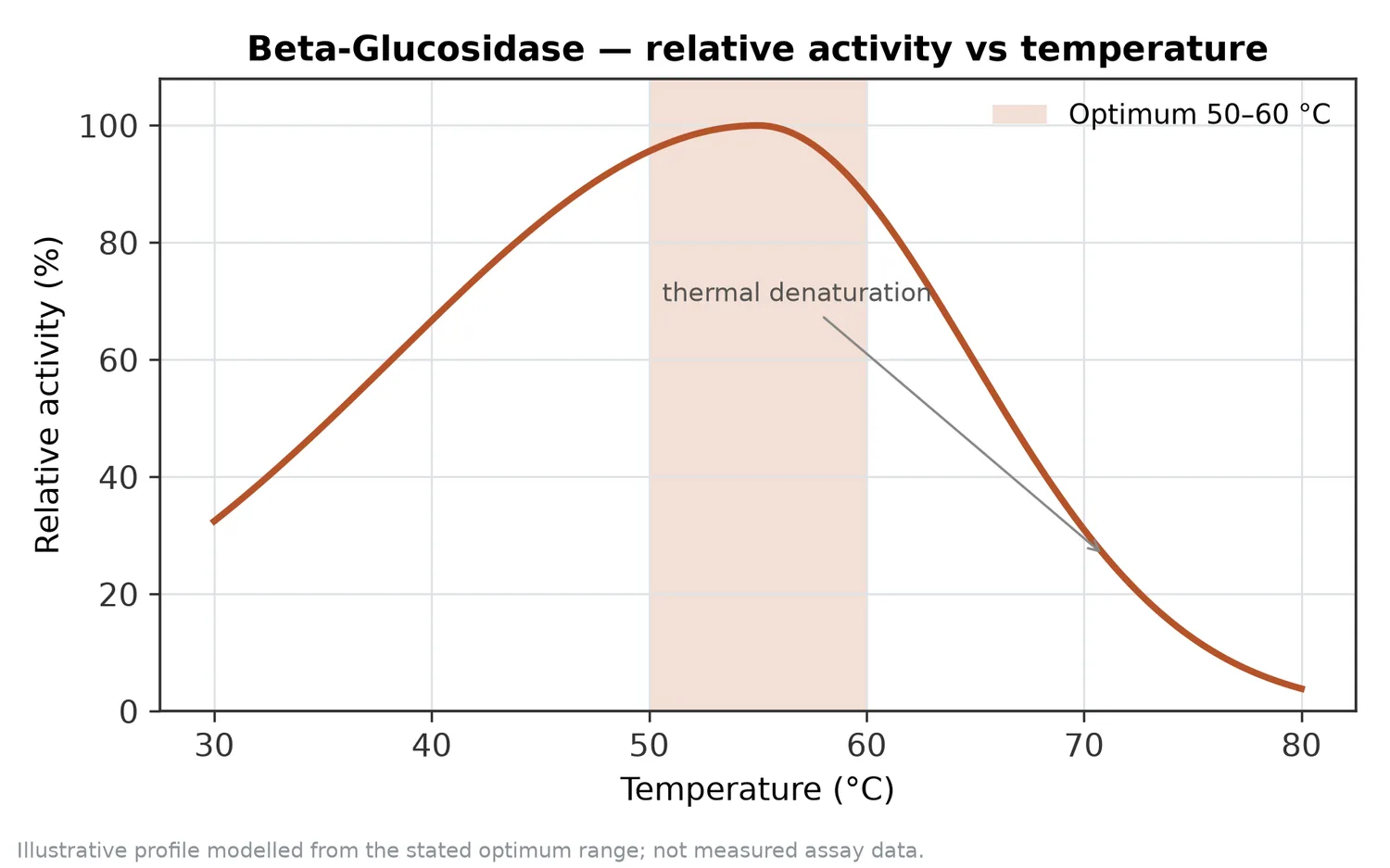
Sıcaklık önemli bir parametredir çünkü enzimatik reaksiyon hızı ile protein stabilitesi arasında denge kurulur. Termostabil beta-glucosidase üzerine yapılan çalışmalar, yüksek sıcaklığa dayanıklı adayların endüstriyel biyokütle dönüşümü ve bazı proseslerde avantaj sağlayabileceğini göstermiştir. Ancak termostabilite, her gıda veya ekstrakt uygulamasında gerekli ya da istenen tek özellik değildir; aroma veya pigment hassasiyeti olan matrislerde daha ılımlı koşullar tercih edilebilir [2].
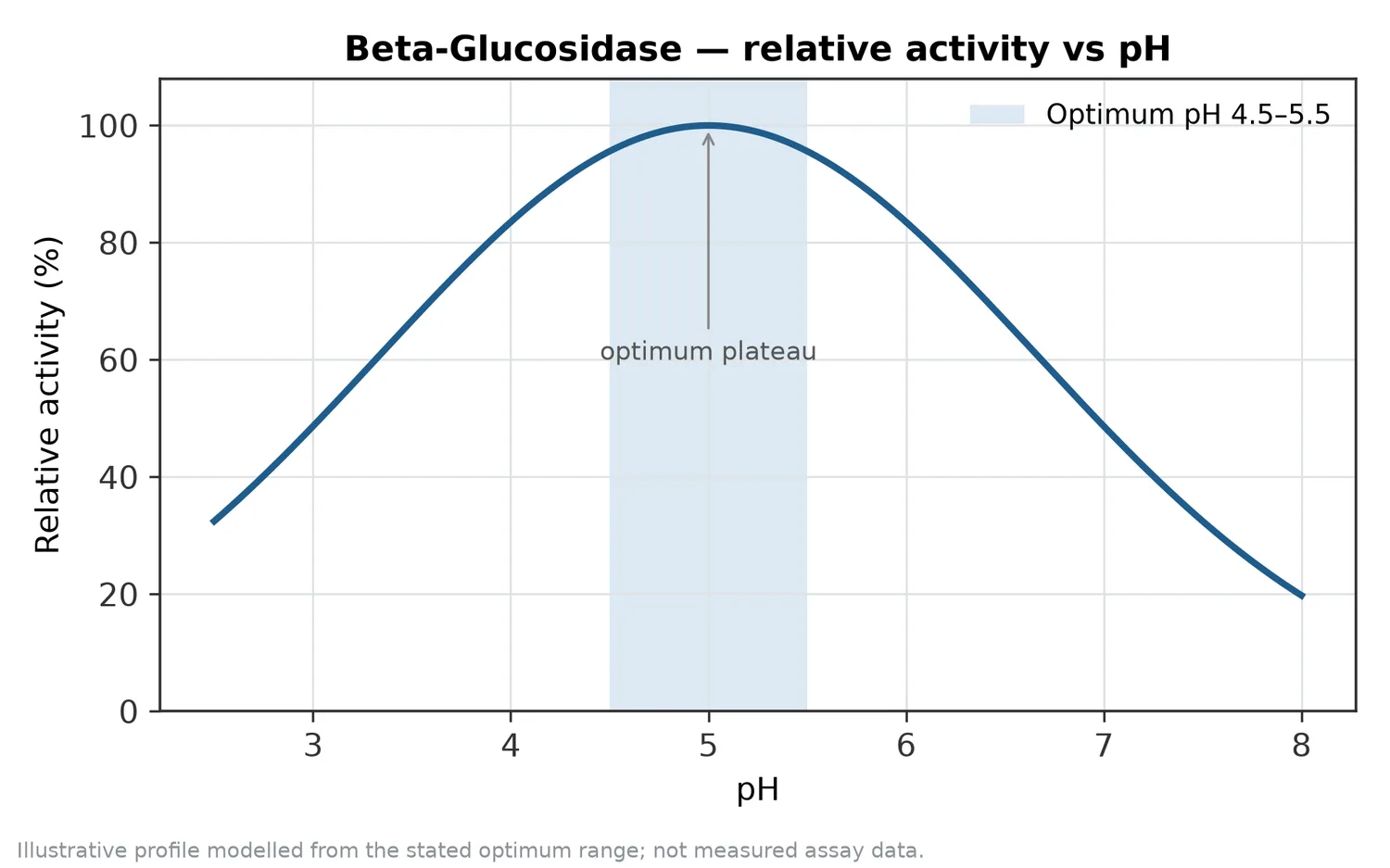
pH da benzer şekilde uygulamaya bağlıdır. Asidik meyve matriksleri, nötre yakın fermentasyon ortamları veya daha farklı endüstriyel prosesler aynı enzime aynı sonucu vermez. Beta-glucosidase’in aktif bölgesindeki iyonlaşabilir gruplar, substrat bağlanması ve suyla hidroliz adımı için uygun protonlanma durumunda olmalıdır; pH bu kimyasal dengeyi doğrudan etkiler [3].
Metal iyonları ve organik çözücü benzeri ortam bileşenleri de aktivite ve stabiliteyi değiştirebilir. Beta-glucosidase nanoflower sistemleri üzerinde metal iyonları ve organik çözeltilerin etkisini inceleyen çalışma, enzimin konformasyonel çevresinin ve immobilizasyon benzeri düzenlemelerin performans üzerinde belirleyici olabileceğini göstermektedir [12].
Glukoz birikimi ise özellikle biyokütle proseslerinde ayrı bir başlıktır. Hidroliz ürünü olan glukoz, bazı beta-glucosidase örneklerinde reaksiyonu yavaşlatan geri besleme etkileri yaratabilir; bu nedenle lignoselülozik sakkarifikasyonda glukoz toleransı veya ürün inhibisyonuna dayanıklılık araştırma konusu olmaya devam eder. Aspergillus niger beta-glucosidase’in sakkarifikasyon ürünleri ve fermantasyon inhibitörlerine tolerans açısından incelenmesi bu pratik gereksinimle ilişkilidir [1].
Endüstriyel kullanımda beta-glucosidase’in yalnızca substratı dönüştürmesi yeterli değildir; proses süresince aktif kalması, matris bileşenlerinden etkilenmemesi ve hedef reaksiyonu sürdürebilmesi gerekir. Bu nedenle literatürde enzim stabilitesi, protein mühendisliği, immobilizasyon ve konformasyon kontrolü gibi konular yoğun biçimde çalışılmıştır [15].
Trichoderma viride beta-glucosidase üzerine yapılan rasyonel moleküler tasarım çalışması, sindirim enzimlerine karşı direnç artırmayı hedeflemiş ve bağlı durum oluşumunun engellenmesi üzerinden dayanıklılık yaklaşımı sunmuştur. Bu tür araştırmalar, beta-glucosidase’in yalnızca endüstriyel reaktörlerde değil, karmaşık biyolojik veya gıda benzeri ortamlarda da davranışının tasarlanabileceğini gösterir [15].
Yüklü peptitlerin beta-glucosidase aktivitesi ve konformasyonu üzerindeki etkisini inceleyen moleküler dinamik çalışması, enzimin çevresindeki elektrostatik ortamın protein hareketliliğini ve katalitik davranışı değiştirebildiğini ortaya koyar. Bu bulgu, formülasyon, taşıyıcı sistem veya immobilizasyon yaklaşımı seçilirken enzimin fiziksel çevresinin dikkate alınması gerektiğini destekler [16].
Beta-glucosidase nanoflower gibi hibrit veya organize yapılarda incelendiğinde, stabilite ve yeniden kullanım potansiyeli öne çıkar. Ancak immobilize veya yapılandırılmış enzim sistemleri serbest enzimle aynı difüzyon davranışına sahip değildir; substratın enzime ulaşması, ürünün ayrılması ve matris viskozitesi performansı etkileyebilir [12].
Beta-glucosidase doğada farklı organizmalarda bulunur; endüstriyel araştırmalarda ise mantarlar, bakteriler, maya sistemleri ve rekombinant ifade platformları öne çıkar. Rekombinant beta-glucosidase’in Escherichia coli içinde saflaştırılması ve karakterizasyonu üzerine çalışma, enzimin laboratuvar ölçeğinde kaynak organizmadan bağımsız olarak üretilebildiğini ve incelenebildiğini gösterir [14].
Bacillus subtilis’te sinyal peptidi optimizasyonuyla termostabil beta-glucosidase salgılanmasının iyileştirilmesi üzerine çalışma, enzimlerin hücre dışına daha verimli taşınmasının biyoproses tasarımında ayrı bir mühendislik problemi olduğunu gösterir. Bu tür çalışmalar tedarik zinciri açısından değil, enzim biyoteknolojisinin nasıl geliştirildiğini anlamak açısından önemlidir [8].
Metagenomik yaklaşım da beta-glucosidase araştırmalarında güçlü bir kaynaktır. Keçi rumen bakterilerinden elde edilen DNA verilerinde beta-glucosidase kodlayan genlerin taranması ve ifade edilmesi, doğal lignoselüloz parçalama ekosistemlerinin yeni enzim adayları için kullanılabileceğini gösterir [17].
Yapısal biyoloji tarafında beta-glucosidase apo yapılarının incelenmesi, substrat bağlanmadan önce enzimin aktif bölge mimarisini anlamaya yardımcı olur. Apo yapı bilgisi, substratla kompleks yapı veya moleküler modelleme çalışmalarıyla birlikte değerlendirildiğinde, bağlanma cebi, esneklik ve katalitik geometri hakkında daha somut yorumlara temel oluşturabilir [18].
Beta-glucosidase, alpha-glucosidase ile aynı şey değildir. Alpha-glucosidase, alfa-glikozidik bağlar üzerinde çalışan ayrı bir enzim grubudur ve özellikle antidiabetik etki mekanizmaları veya karbonhidrat sindirimi bağlamında çok farklı bir literatüre sahiptir. Sarımsak gibi bitkisel ürünlerde alpha-glucosidase inhibisyonunun incelendiği çalışmalar, beta-glucosidase uygulamalarından ayrı değerlendirilmelidir [19].
Beta-glucosidase, beta-glucan ile de karıştırılmamalıdır. Beta-glucan bir gıda lifi veya yapısal polisakkarit olarak ele alınır; kaynağı, ekstraksiyonu, çözünürlüğü, viskozite etkisi ve sağlıkla ilişkilendirilen fonksiyonları ayrı bir konu başlığıdır. Beta-glucosidase ise belirli beta-glukozit bağlarını hidroliz eden enzimdir; beta-glucan literatüründeki lif etkileri doğrudan beta-glucosidase ürün iddiası olarak yorumlanmamalıdır [20].
Bir diğer karışıklık, beta-glucosidase’in her glikozit yapıyı aynı hızda ve aynı verimle dönüştüreceği varsayımıdır. Enzim-substrat uyumu; glukozun bağlandığı aglikonun hacmi, polaritesi, sterik engeli ve bağ çevresindeki kimyasal yapı tarafından belirlenir. Botryodiplodia theobromae beta-glucosidase üzerine yapılan mekanizma ve kinetik çalışmalar, farklı substratların aynı enzime farklı davranış gösterebileceğini ortaya koyan erken örneklerdendir [21].
Beta-glucosidase için en güçlü kanıt alanı, glukozit bağının hidrolizi ve sellobiyoz benzeri ara ürünlerin glukoza dönüştürülmesidir. Bu alan hem mekanistik çalışmalarla hem de lignoselülozik biyokütle sakkarifikasyonu bağlamındaki uygulama çalışmalarıyla desteklenir [2].
Biyokütle ve biyorefinery uygulamaları da güçlü teknik temele sahiptir; çünkü selüloz hidroliz zincirinde beta-glucosidase’in görevi net biçimde tanımlanabilir. Aspergillus niger beta-glucosidase’in lignoselülozik sakkarifikasyon ürünleri ve fermantasyon inhibitörlerine tolerans açısından incelenmesi, gerçek proses ortamlarının enzim performansını nasıl sınayabileceğini gösterir [1].
Gıda, içecek ve bitkisel ekstrakt uygulamaları ise daha matris bağımlıdır. Glukozitlerin enzimatik hidrolizi kimyasal olarak açıktır; ancak aroma yoğunluğu, renk stabilitesi, fenolik profil, biyolojik erişilebilirlik veya duyusal kalite gibi sonuçlar hammaddenin bileşimine ve proses koşullarına bağlıdır. Bitkisel biyoaktif bileşenler üzerine yapılan pancar ve deniz pancarı derlemeleri, hammadde kompozisyonunun ürün etkisini belirleyen çok değişkenli bir konu olduğunu gösterir [22].
Biyokatalitik sentez ve transglikozilasyon gibi alanlar da dikkatli yorumlanmalıdır. Beta-glucosidase belirli koşullarda glukoz transferi yapabilir; ancak suyun bol olduğu ortamlarda hidroliz çoğu zaman baskın olabilir. Mammalian cytosolic beta-glucosidase üzerinde transglikozilasyonun mekanizma probu olarak kullanılması, bu reaksiyonun mümkün olduğunu fakat koşul bağımlı değerlendirilmesi gerektiğini gösterir [7].
Enzymes.bio, Beta-Glucosidase ürününü çevrim içi satışa sunan bir tedarikçidir; üretici, analiz laboratuvarı veya uygulama testi sağlayıcısı olarak konumlandırılmaz. Ürün 1 kg birimler halinde doğrudan satın alınır; çevrim içi ödeme tamamlandıktan sonra sipariş işleme alınır ve teslimat süreci başlatılır.
Siparişle birlikte Analiz Sertifikası (CoA) ve Güvenlik Bilgi Formu (SDS) sağlanır. Bu dokümanlar, ürünün ilgili parti bilgileri ve güvenli kullanım çerçevesi için kullanılır; uygulama performansı ise hedef substrat, proses matriksi, sıcaklık, pH, temas süresi ve diğer formülasyon bileşenlerine bağlı olarak değişebilir.
Beta-glucosidase, doğru proses bağlamında güçlü ve çok yönlü bir biyokatalizördür: lignoselülozik biyokütlede sellobiyozun glukoza ilerletilmesi, bitkisel glikozitlerin dönüştürülmesi ve belirli koşullarda transglikozilasyon potansiyeli aynı temel kimyasal kabiliyetten doğar. Güvenilir kullanım yaklaşımı, enzimi genel bir “her şeyi dönüştüren katkı” olarak değil, beta-glikozidik bağ taşıyan hedef substratlara yönelik seçici bir proses aracı olarak değerlendirmektir [23].
1 kg birimler halinde satılır; stokta mevcut ve sevkiyata hazırdır. Mağazamızdan doğrudan sipariş verin — online ödeme yapın, siparişinizi işleme alalım. Her siparişe Analiz Sertifikası ve Güvenlik Bilgi Formu dahildir.
Beta-Glucosidase satın alın →İlk atıf sırasına göre numaralandırılmıştır. Açık erişimli kaynaklardır; her birinin yayım sırasında erişilebilir olduğu doğrulanmıştır. Metindeki atıf numaraları buraya bağlantı verir.