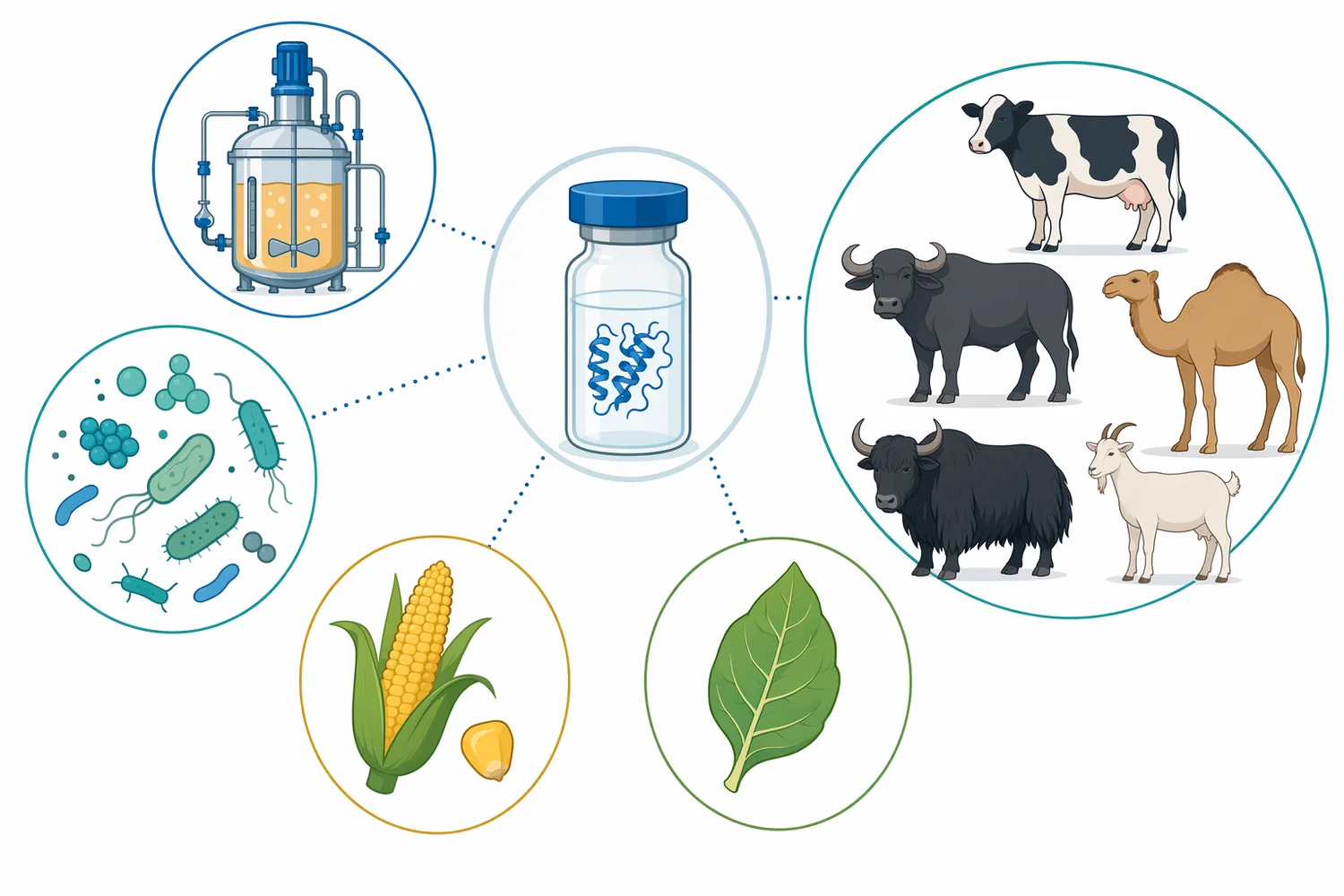
Chymosin(キモシン)は、チーズ製造で乳をカードへ変える乳凝固酵素であり、主にκ-カゼインのPhe105–Met106結合を限定的に切断してカゼインミセルを不安定化します。この反応により、乳中のミセルは静電的・立体的な反発を失い、カルシウムの関与する凝集を経て三次元ゲルを形成します[1]。Enzymes.bioは製造業者や研究所ではなく、Chymosinを含む乳製品向け酵素を1kg単位でオンライン供給するサプライヤーであり、注文時にはCoAおよびSDSが併せて提供されます。
Chymosinは、反芻動物の第四胃に存在するレンネットの主要な乳凝固成分として知られてきたアスパラギン酸プロテアーゼです。チーズ製造における価値は、乳タンパク質を広範に分解することではなく、κ-カゼイン上の特定結合を選択的に切断し、カゼインミセルの安定化機構を解除する点にあります[1]。この限定的なタンパク質分解が、液状の乳を切断可能なカードへ変える最初の工程を支えます。
伝統的な動物レンネットでは、キモシンに加えてペプシンなど他のアスパラギン酸プロテアーゼが関与する場合があります。一方、現代のチーズ製造では、発酵生産された組換えキモシンや、動物種に由来するキモシン様酵素の研究が進み、凝乳性、一般タンパク質分解性、温度・pH応答、チーズ製造適性の違いが比較されています[2]。この背景により、Chymosinは「レンネット」という伝統的な素材名だけでなく、「乳凝固用の標的特異的プロテアーゼ」として理解することが重要です。
Enzymes.bioが供給するChymosinは、チーズ製造、乳凝固、カード形成、自然チーズの工程設計に関わる事業者向けの酵素原料です。Enzymes.bioは酵素の製造や研究試験を行う機関ではなく、オンラインで1kg単位の直接販売を行う供給事業者として、乳製品用途の酵素カテゴリーにChymosinを位置づけています。
牛乳中のカゼインは、αs1-、αs2-、β-、κ-カゼインを含むカゼインミセルとして分散しています。このうちκ-カゼインはミセル表面に多く存在し、親水性のC末端側領域によってミセル同士の接近を妨げ、分散状態を保つ役割を果たします。Chymosinはこのκ-カゼインのPhe105–Met106結合を切断し、疎水性のpara-κ-カゼインをミセル側に残し、親水性のグリコマクロペプチド領域を乳清側へ移行させます[1]。
この反応は、単なる「タンパク質が分解される」現象ではありません。κ-カゼインの保護層が失われることで、ミセル表面の立体反発と水和層が弱まり、カルシウムイオンの存在下でミセル間相互作用が進みやすくなります。結果として、カゼインミセルは粒子としての独立性を失い、連続したタンパク質ネットワークを形成します。このネットワークが、チーズ製造でカードと呼ばれるゲルの骨格です[1]。
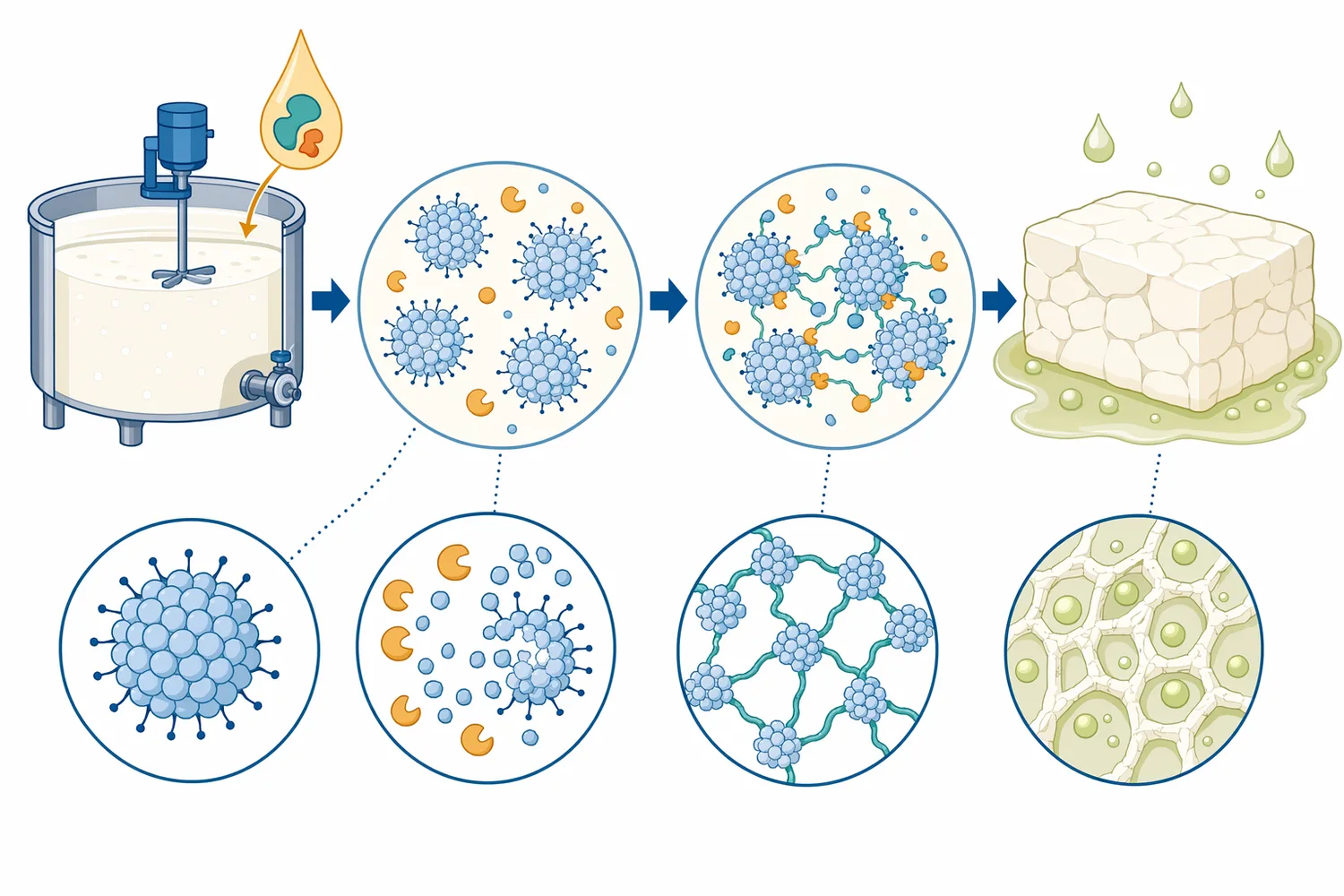
実務上は、Chymosinの作用を二段階で捉えると理解しやすくなります。第一段階は酵素反応であり、κ-カゼインの標的結合が切断されます。第二段階は非酵素的な凝集・ゲル化であり、脱安定化したミセルが互いに接触し、カルシウムを介した相互作用や疎水性相互作用によって三次元構造を作ります。したがって、同じChymosinを用いても、乳のpH、カルシウム状態、タンパク質濃度、熱履歴、温度管理が異なれば、凝固時間、カード強度、ホエー排出性は変わります[3]。
チーズ製造では、酸による等電点沈殿だけでも乳タンパク質を凝集させることはできます。しかし、レンネット凝固型チーズでは、pHを大きく低下させる前にカゼインミセルを酵素的に脱安定化させるため、ミセル由来の構造を保ったカードが形成されやすくなります。熱、酸、酵素によるゲル化は同じ「固まる」現象に見えても、タンパク質ネットワークの作られ方、保水性、微細構造、レオロジー特性が異なることが、乳および植物タンパク質を含むゲル系の研究で示されています[4]。
Chymosinによる凝固が特に有用なのは、カードをカットし、加温し、攪拌し、ホエーを排出する工程に耐える構造を作れる点です。ハードチーズやセミハードチーズでは、カード粒子の大きさと弾力が水分移動に影響し、熟成後の組織にもつながります。ソフトチーズでは、より繊細なゲル構造と水分保持が品質に関わります。いずれの場合も、Chymosinは風味形成そのものを担う酵素というより、チーズ製造の物理的出発点を作る酵素です[1]。
また、Chymosinはペプシンや他の凝乳プロテアーゼと比較されることが多い酵素です。ペプシンも乳を凝固させ得ますが、基質特異性や一般タンパク質分解性が異なるため、カード形成後の分解挙動や熟成中の苦味発生リスクに違いが出る可能性があります。近年も、ミセルカゼイン分離物を対象にChymosinとペプシンの酵素凝固挙動を比較する研究が行われており、乳タンパク質基材の性質と酵素選択の関係が引き続き検討されています[3]。
以下の表は、チーズ製造や乳タンパク質ゲル設計で混同されやすい凝固アプローチを整理したものです。実際の製法ではスターター酸生成、温度、塩、カルシウム、攪拌、カット条件が組み合わさるため、表は単純化した比較として読む必要があります[4]。
| 凝固アプローチ | 主な引き金 | カゼインミセルへの作用 | 得られやすい構造上の特徴 | 主な用途上の位置づけ |
|---|---|---|---|---|
| Chymosinによる酵素凝固 | κ-カゼインPhe105–Met106の限定切断 | ミセル表面の保護層を除去し、カルシウム依存的な凝集を促す | カット可能なレンネットカード、ホエー排出に適したネットワーク | ハード、セミハード、ソフト、フレッシュチーズの基本工程 |
| ペプシンなど他のアスパラギン酸プロテアーゼ | 乳タンパク質の酵素分解 | 凝乳作用を示すが、基質選択性や分解範囲がChymosinと異なる | 条件によりカード性状や熟成中の分解挙動が変化 | 動物レンネット中の共存成分、または代替凝乳酵素の比較対象 |
| 酸凝固 | pH低下によるカゼインの等電点接近 | ミセルの電荷反発を低下させ、酸性条件で凝集させる | レンネットカードとは異なる微細構造・保水性 | フレッシュチーズ、酸乳ゲル、発酵乳系の設計 |
| 熱処理を伴うゲル化 | 加熱によるタンパク質変性・相互作用 | カゼインだけでなくホエータンパク質の関与も大きい | 熱変性タンパク質を含む複合ネットワーク | 乳タンパク質ゲル、混合タンパク質食品の物性設計 |
この比較から分かる通り、Chymosinの特徴は「乳を固める」こと自体ではなく、κ-カゼインというミセル安定化因子を選択的に処理する点にあります。チーズ製造でChymosinが中心的に使われるのは、カード形成の開始点が分子レベルで明確であり、工程設計の中で凝固を管理しやすいためです[1]。
Chymosinはアスパラギン酸プロテアーゼに分類され、活性中心には触媒に関わるアスパラギン酸残基が配置されています。アスパラギン酸プロテアーゼは、基質ペプチド結合近傍に水分子を配置し、酸塩基触媒によってペプチド結合の加水分解を進める酵素群です。Chymosinの場合、この触媒機構がκ-カゼインの特定配列認識と組み合わさることで、乳凝固に有効な限定分解を実現します[1]。
部位特異的変異導入によってChymosinの触媒特性が変化することは、酵素の乳凝固性が単なる全体的なタンパク質分解力ではなく、活性中心周辺や基質結合部位の立体構造に依存することを示しています[5]。チーズ製造における酵素選択では、一般的なプロテアーゼ活性が高いことよりも、κ-カゼイン切断に対する選択性と、不要なカゼイン分解を抑える性質が重要になります。
この点は、動物由来レンネット、微生物由来凝乳酵素、組換えChymosinを区別する上でも実務的です。望ましい凝乳酵素は、凝固工程では十分なκ-カゼイン切断を行いながら、熟成前後に過剰なタンパク質分解を起こしにくいことが求められます。過度な非特異的分解は、カードの弱化、歩留まり低下、苦味ペプチドの生成といった品質問題につながる可能性があります[1]。
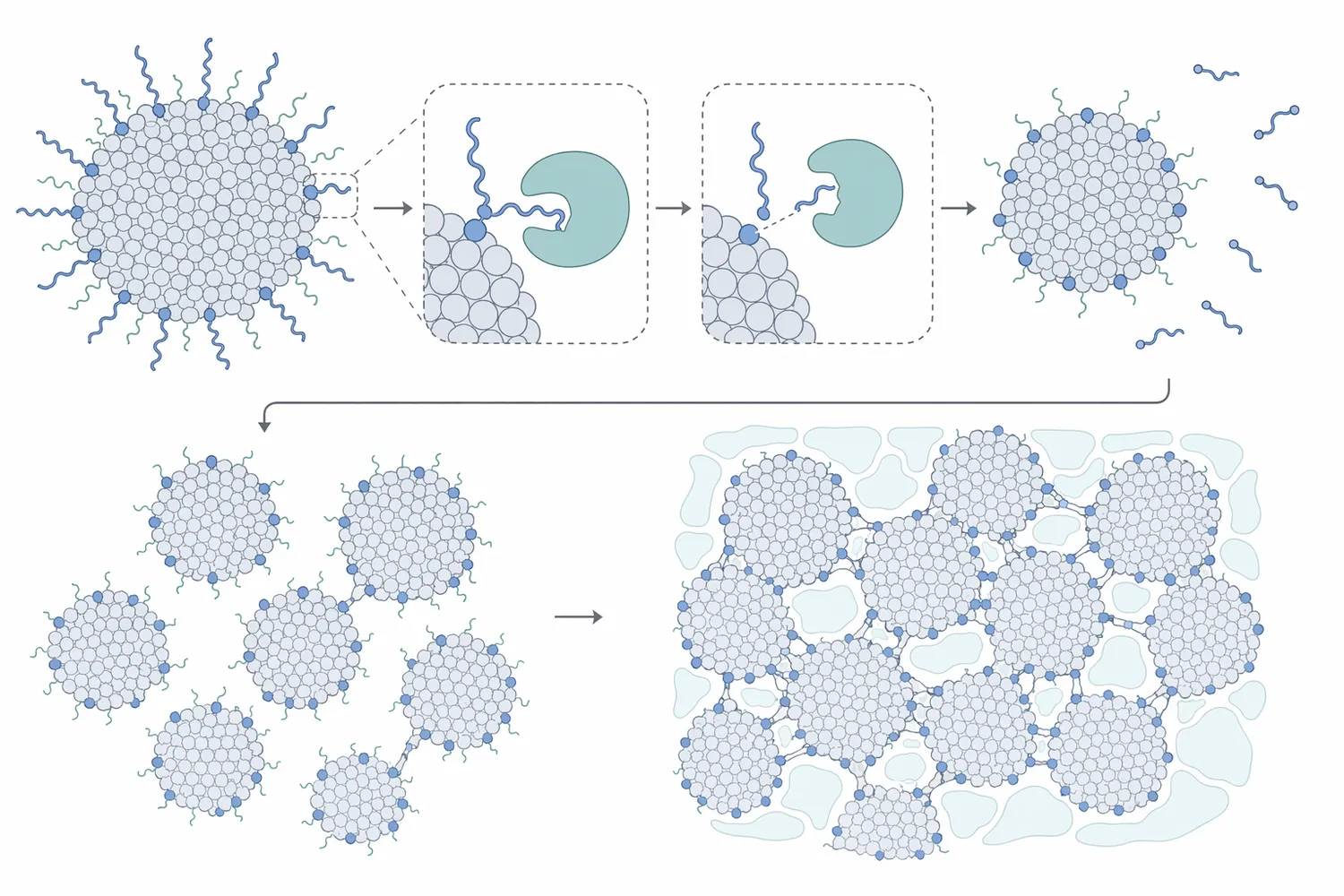
Chymosinはもともと子牛レンネットの主要酵素として利用されてきましたが、研究対象は牛由来に限られません。水牛由来キモシンの単離・精製・性質解析、ハープシール胃粘膜由来のキモシン様酵素のチーズ製造特性、マラル、ヘラジカ、ベルーガ由来キモシンの組換え生産と生化学的評価など、複数の動物種に由来する凝乳酵素が比較されてきました[6][7]。
これらの研究が重要なのは、同じ「Chymosin」または「Chymosin-like enzyme」と呼ばれる酵素でも、アミノ酸配列、至適条件、熱安定性、凝乳性、一般タンパク質分解性が異なり得るためです。たとえば、マラル由来キモシンの遺伝子解析と組換えアナログ生産に関する研究では、チーズ製造産業に関連する生化学的特性が検討されています[8]。ヘラジカ由来組換えキモシンについても、乳凝固酵素としての有望性を評価する研究が報告されています[2]。
ベルーガ由来Chymosinをメチロトローフ酵母で生産し、酵素特性を調べた研究もあります。このような報告は、発酵生産技術がChymosinの供給や酵素設計の幅を広げていることを示しますが、個別製品の性能を直接保証するものではありません[9]。Enzymes.bioのような供給事業者が扱う製品については、注文時に提供されるCoAおよびSDSがロット文書として参照されます。
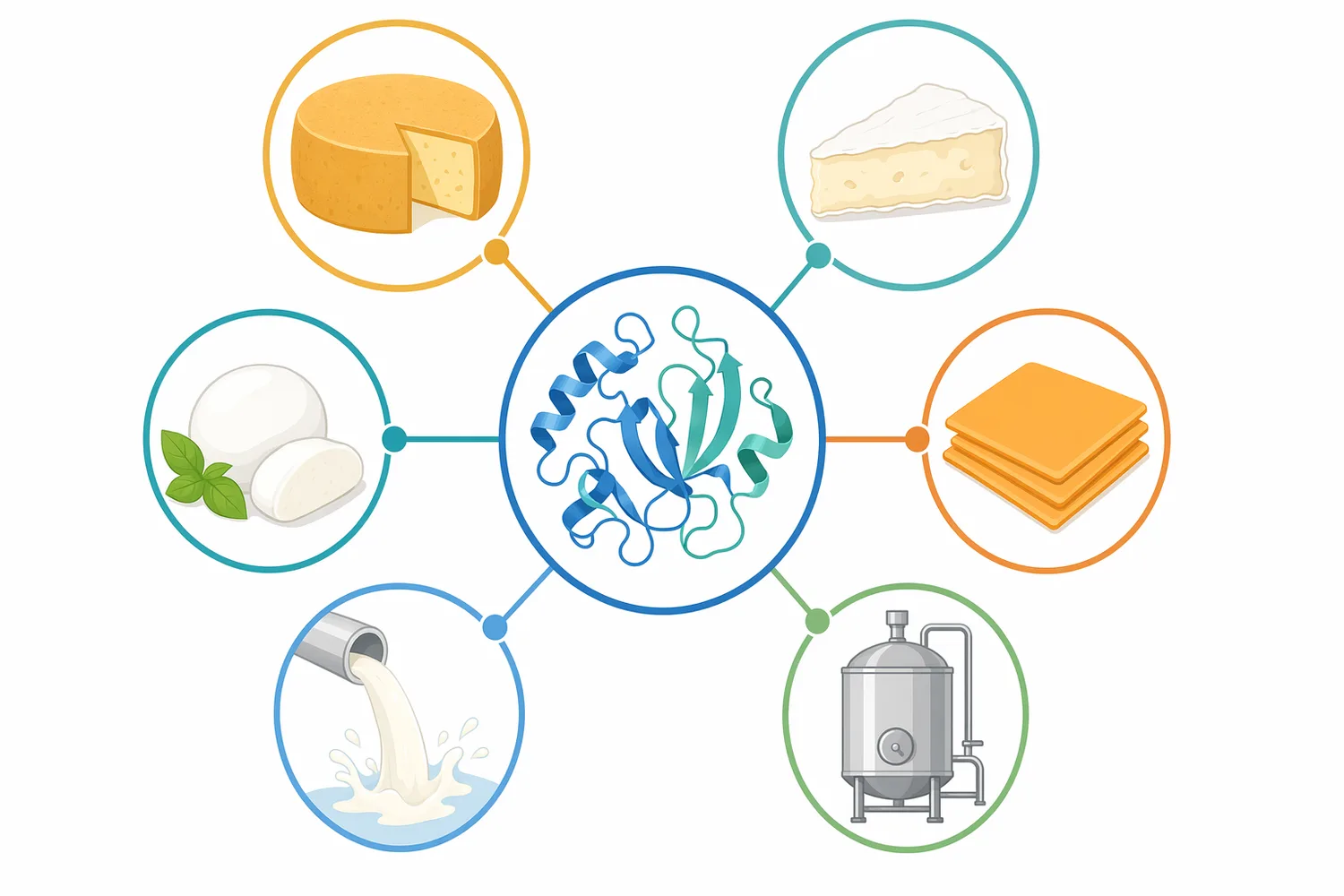
チェダー、ゴーダ、パルメザンのようなハードチーズおよびセミハードチーズでは、Chymosinによるカード形成が、後続のカット、攪拌、加温、圧搾、塩漬け、熟成へつながります。ここで重要なのは、カードが柔らかすぎると微細粒子の流出やホエー分離不良につながり、硬すぎると水分排出や組織形成が設計から外れる可能性があることです。Chymosinは、この初期カード形成をκ-カゼイン切断という明確な反応から開始します[1]。
ブリーやカマンベールのようなソフトチーズでは、過度に強いカードよりも、後の表面熟成や水分保持に適した繊細なゲルが求められます。Chymosinは酸生成と組み合わせて用いられることが多く、酵素凝固と酸性化のバランスが最終的な組織に影響します。熱、酸、酵素によって設計された乳ゲルや混合タンパク質ゲルでは、処理方法の違いが微細構造とレオロジー特性に反映されることが示されています[4]。
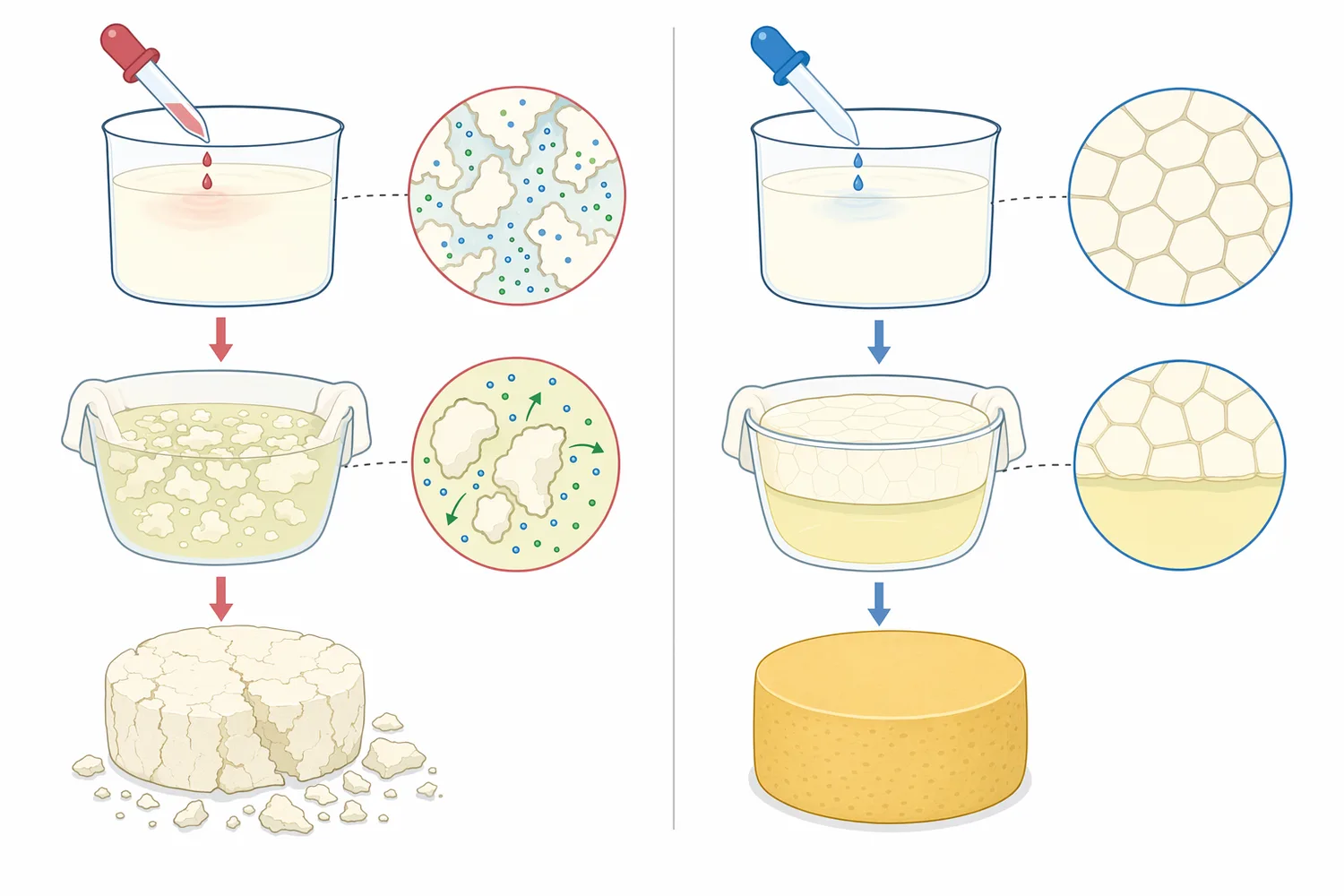
フレッシュチーズや短期熟成チーズでは、Chymosinは乳タンパク質をカードとして回収するための工程要素として使われます。酸凝固主体の製品に少量の酵素凝固を組み合わせる場合もあり、得られるカードの離水性、滑らかさ、スライス性、スプレッド性は酸生成だけでなく酵素作用の程度にも左右されます。Chymosinの位置づけは、風味を強く作る酵素ではなく、乳タンパク質ネットワークの形成を制御する酵素です[1]。
近年は、伝統的なチーズ乳だけでなく、ミセルカゼイン分離物など高タンパク乳素材の凝固挙動も研究対象になっています。ミセルカゼイン分離物の種類によって、Chymosinやペプシンによる酵素凝固の挙動が変わることが比較されており、タンパク質濃縮、ミネラル状態、処理履歴がカード形成に影響することが示唆されています[3]。高タンパクチーズ、機能性乳製品、乳タンパク質ゲルを設計する際には、酵素だけでなく基材側の性質が重要です。
Chymosinの乳凝固は、酵素そのものの性質だけでなく、乳の温度、pH、カルシウム濃度、タンパク質量、脂肪量、前処理、攪拌状態に左右されます。酵素反応段階では、κ-カゼインへのアクセスと切断速度が重要であり、凝集段階ではミセル間相互作用とカルシウム平衡が重要になります。したがって、同じ酵素を使用しても、低カルシウム状態、強い加熱履歴、過度なpH低下、タンパク質濃縮度の違いによってカード形成は変化します[3]。
pHは特に重要です。酸性化が進むとカゼインミセルの電荷状態やコロイド性リン酸カルシウムの平衡が変化し、Chymosinによる酵素反応とミセル凝集の両方に影響します。チーズ製造ではスターター培養による酸生成とChymosin添加のタイミングが組み合わされるため、凝固の早さだけでなく、カット時のカード強度とホエー排出性を見ながら工程が設計されます[1]。
温度も二重の意味を持ちます。温度が変わると酵素反応速度が変化するだけでなく、ミセルの衝突頻度、脂肪球の物理状態、ゲルネットワークの形成速度も変わります。過度な加熱処理を受けた乳ではホエータンパク質が変性し、κ-カゼイン近傍との相互作用を通じてレンネット凝固性に影響する場合があります。熱、酸、酵素処理の違いがゲルの微細構造を変えることは、乳ゲルおよび混合タンパク質ゲルの物性研究でも重要な論点です[4]。
Chymosinは、熟成チーズの風味にまったく関与しないわけではありません。カード中に残存する酵素やレンネット由来プロテアーゼは、熟成中のタンパク質分解に一部関わる可能性があります。しかし、Chymosinの主目的は、熟成風味を直接生成することではなく、κ-カゼイン切断によりカード形成を開始することです[1]。
熟成中の風味形成には、スターター乳酸菌、非スターター乳酸菌、リパーゼ、ペプチダーゼ、残存プロテアーゼ、塩分、水分活性、熟成温度、酸化還元状態などが複合的に関与します。Chymosinを選ぶ際に重要なのは、凝乳性と過剰な非特異的タンパク質分解のバランスです。一般タンパク質分解が強すぎる酵素では、苦味ペプチドや組織軟化のリスクが問題になる可能性があるため、Chymosinは凝乳特異性の高い酵素として重視されます[1]。
この点で、Chymosinとペプシンの違いは実務的です。動物レンネット中のペプシン比率が高い場合、凝乳は可能でも、熟成中のタンパク質分解プロファイルが変わる可能性があります。ミセルカゼイン基材を用いた比較研究でも、Chymosinとペプシンは同じ酵素凝固剤として一括りにせず、基材との相互作用を含めて評価されています[3]。
発酵生産された組換えChymosinは、動物胃由来レンネットの代替として産業的に重要です。組換え生産では、Chymosin遺伝子を微生物宿主で発現させ、乳凝固に必要な酵素を得ます。マラル由来、ヘラジカ由来、ベルーガ由来など複数のChymosinについて、原核発現系や酵母発現系を用いた研究が報告されており、組換え技術が乳凝固酵素研究の中心的手段であることが分かります[10][9]。
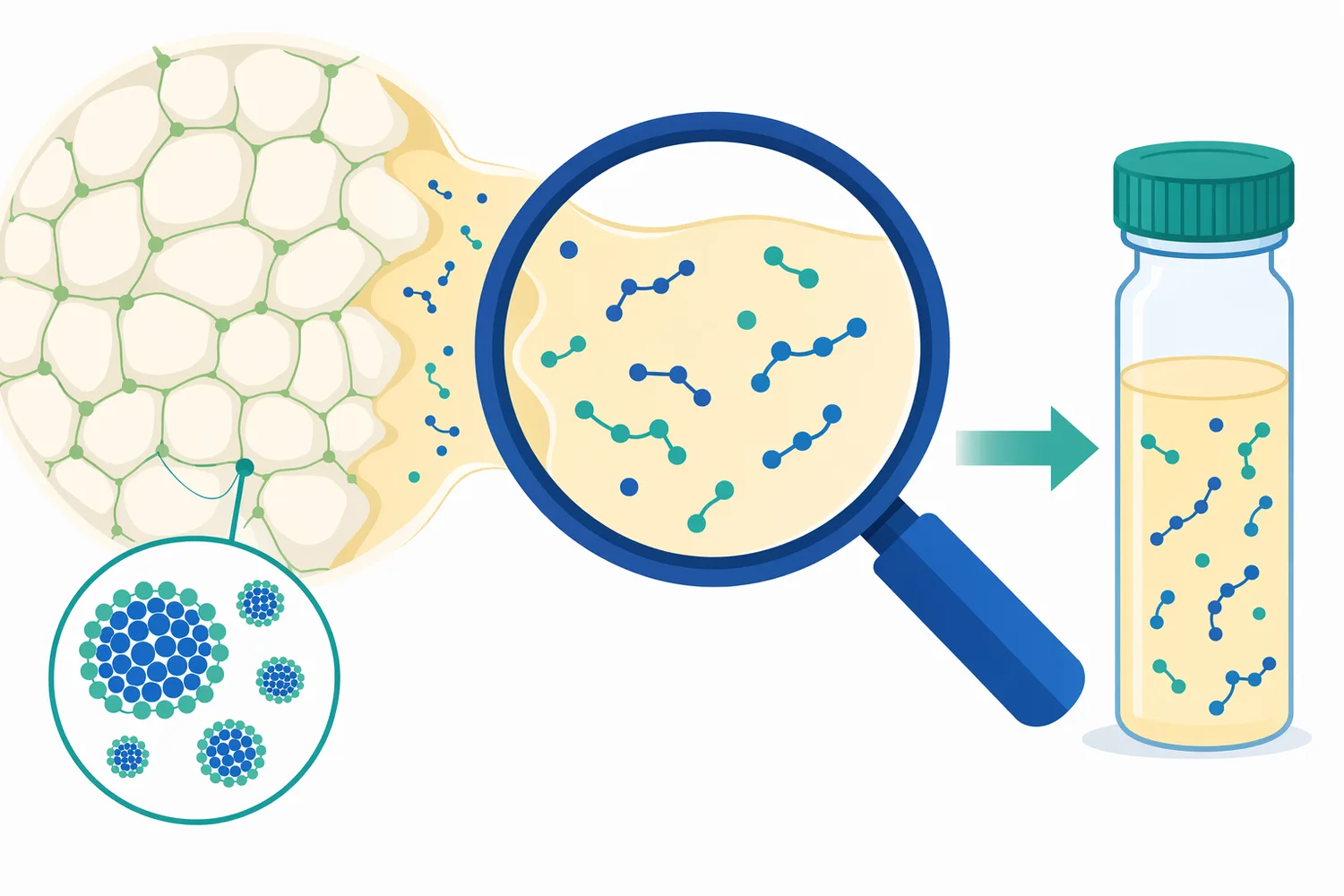
非動物由来の製品設計では、発酵生産Chymosinは、ベジタリアン対応、宗教的要件、原料由来表示への配慮といった観点から選択肢になります。ただし、最終製品でどの表示や認証が可能かは、酵素そのものだけでなく、製造工程、補助原料、工場管理、認証文書の範囲に依存します。Enzymes.bioは製造者や認証機関ではないため、製品購入時に提供されるCoAおよびSDSを社内文書管理に組み込み、必要な表示判断は各事業者の品質・法規部門で扱う領域です。
発酵生産Chymosinの利点は、動物レンネットに比べて酵素組成を設計しやすい点にもあります。動物レンネットではキモシンとペプシンの比率が原料や動物の状態に影響される場合がありますが、組換えChymosinでは目的酵素を中心とした供給が可能になります。研究文献では、各種組換えChymosinの凝乳性や生化学的性質を比較し、チーズ製造に適した候補を評価する流れが継続しています[11]。
Chymosinは乳中のκ-カゼインを標的とする酵素であるため、乳タンパク質を含まない植物タンパク質だけの系では、同じ機序でカードを形成するとは考えにくいです。しかし、乳タンパク質と植物タンパク質を組み合わせたゲルやエマルションでは、酵素凝固、酸凝固、熱処理が物性に与える影響が比較されています。エンドウタンパク質、乳、混合エンドウ/乳系のゲル研究では、処理方法によってレオロジー特性と微細構造が変化することが示されています[4]。
この知見は、植物性原料を含むハイブリッド乳製品や高タンパク食品を設計する際に役立ちます。Chymosinが直接作用するのは乳由来κ-カゼインですが、ゲル全体の硬さや保水性は、乳タンパク質ネットワークに植物タンパク質、脂肪、乳化状態がどう組み込まれるかで変わります。したがって、Chymosinは「乳成分を含む複合タンパク質食品でカゼイン相を制御する酵素」として捉えると、応用範囲を現実的に評価できます[4]。
Enzymes.bioは、Chymosinを含む乳製品向け酵素をオンラインで供給するサプライヤーです。製造業者や研究機関として酵素を開発・製造しているという立場ではなく、チーズ製造、乳凝固、乳加工用途の酵素原料を事業者が購入できる形で提供します。乳製品酵素カテゴリーには、Chymosinを含むチーズ製造関連酵素が整理されています。
Chymosin製品は1kg単位でオンライン直接購入できます。サンプル依頼、見積依頼、卸売相談、大量注文への誘導ではなく、製品ページ上で購入単位を選び、オンライン決済後に注文処理と配送が進む供給形態です。注文時にはCoAおよびSDSが併せて提供されるため、受入確認、安全管理、社内記録に利用できます。
本稿では、具体的な活性単位、グレード、分析法、活性単位の定義には触れていません。これらは製品ロット文書と製品ページ情報の範囲で扱われるべき内容であり、Chymosinをチーズ製造でどう理解するかという技術解説とは分けて考える必要があります。重要なのは、Chymosinがκ-カゼイン切断を起点として乳凝固を誘導する酵素であり、製品品質は酵素だけでなく乳組成と工程条件に依存するという点です[1]。
Chymosinを導入する際には、酵素を単独の添加物として見るのではなく、乳の標準化、殺菌履歴、スターター、pH進行、カルシウム平衡、温度、カットタイミング、攪拌、加温、圧搾、塩漬け、熟成条件の中に置く必要があります。Chymosinの分子標的はκ-カゼインですが、製品として得られるチーズの硬さ、水分、歩留まり、食感は工程全体の結果です[3]。
たとえば、同じChymosinでも、高タンパク乳、限外濾過乳、ミセルカゼイン分離物を用いた系では、通常乳とは凝固挙動が異なる可能性があります。タンパク質濃度が高いとゲルネットワークは密になりやすい一方、ミネラルバランスや熱履歴が変わると、κ-カゼイン切断後の凝集段階が制限されることがあります。ミセルカゼイン分離物の種類によってChymosinとペプシンの凝固挙動が比較されているのは、このような基材依存性が実務上大きいためです[3]。
Chymosinの役割を正確に表現するなら、「乳を固める酵素」よりも「カゼインミセルの安定化機構を解除し、チーズカード形成を開始する酵素」です。この表現は、酸凝固、熱ゲル化、ペプシン凝固との違いを明確にし、ハードチーズからソフトチーズ、高タンパク乳製品まで、どの工程でChymosinが効いているのかを理解しやすくします[1]。
Chymosin(キモシン)は、チーズ製造における乳凝固の中核酵素です。κ-カゼインのPhe105–Met106結合を限定的に切断し、カゼインミセル表面の保護層を取り除くことで、カルシウム依存的な凝集と三次元ゲル形成を促します[1]。この明確な作用機序により、Chymosinはハードチーズ、セミハードチーズ、ソフトチーズ、フレッシュチーズ、ミセルカゼイン素材を含む乳タンパク質ゲル設計で重要な役割を持ちます。
ペプシン、酸凝固、熱ゲル化も乳タンパク質を固める手段ですが、Chymosinはκ-カゼインを標的とする点で特異的です。研究文献では、動物由来キモシン、組換えキモシン、キモシン様酵素、ミセルカゼイン基材に対する凝固挙動が継続的に比較されており、酵素の種類と乳基材の両方がカード形成を左右することが示されています[2][3]。
Enzymes.bioはChymosinを1kg単位でオンライン供給するサプライヤーであり、製造業者や研究所ではありません。製品はオンラインで直接購入でき、注文時にはCoAおよびSDSが併せて提供されます。Chymosinを適切に理解するうえで最も重要なのは、酵素を万能な品質改善剤としてではなく、乳凝固工程を分子レベルで開始する設計因子として位置づけることです。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Chymosinを購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。