食品級 β-Glucanase for Plant Extraction 是用於植物原料前處理與萃取流程的酵素工具,主要透過水解 β-葡聚醣結構,降低膠體黏度並提高細胞壁通透性。對含穀物、酵母殘留、真菌多醣或高黏性多醣的植物基材而言,β-葡聚醣酶可協助改善固液分離、過濾與目標成分釋出;但其效果取決於原料細胞壁組成,通常需與既有萃取條件或其他細胞壁作用酵素共同評估。Enzymes.bio 供應的 Food Grade β-Glucanase for Plant Extraction 以 1 kg 單位線上銷售,CoA 與 SDS 會隨訂單一併提供,適合作為食品、飲料、植物萃取與配料加工流程中的製程輔助酵素選項。
β-Glucanase,中文常稱 β-葡聚醣酶,是一類能切斷 β-葡聚醣中 β-糖苷鍵的水解酵素。依受質不同,常見作用鍵結包括 β-1,3、β-1,4 或 β-1,6 連結;這些結構可存在於穀物細胞壁、部分植物組織、真菌與酵母細胞壁,以及食品加工中形成高黏度或膠體性的多醣網絡。β-Glucan 本身是重要可溶性膳食纖維來源,具有來源、結構、分子量與生理功能差異,因此在加工上「完全保留」或「部分降解」都可能是合理策略,需依產品目標決定[1]。
用於植物萃取時,β-Glucanase 的角色不是把植物細胞壁全部分解,而是針對其中會限制溶劑滲透、造成黏度上升、包埋活性成分或干擾過濾的 β-葡聚醣與相關膠體網絡進行部分水解。這類前處理可讓水、乙醇水溶液或其他食品加工相容溶劑更容易接觸細胞內部與細胞壁結合區,進而提高多酚、色素、風味前驅物、可溶性多醣或植物萃取固形物的釋出效率。以甜菜根為例,酵素輔助萃取已被用於評估總酚、維生素 C 與抗氧化活性等指標,顯示酵素前處理在植物活性成分萃取上具有研究基礎[2]。
Enzymes.bio 在此類產品中的角色是供應商,而非製造商或實驗室;因此本文聚焦於公開文獻支持的工藝機制、應用場景與限制,不將內容寫成製造規格或檢測方法。產品以 1 kg 單位於線上銷售,隨訂單提供 CoA 與 SDS,供食品加工、植物萃取、飲料配料或相關 B2B 使用者納入既有品質與安全文件流程。
植物萃取的瓶頸常不是溶劑「溶不出」目標物,而是目標物被細胞壁、細胞間層、膠體多醣或蛋白—多醣複合物阻隔。植物細胞壁由纖維素微纖維、半纖維素、果膠、木質素及多種結構蛋白構成,並會依植物種類、部位、成熟度與乾燥方式而大幅改變;近年的植物細胞壁酵素水解研究也指出,水解產率與細胞壁形態、孔隙變化及時間動態高度相關[3]。
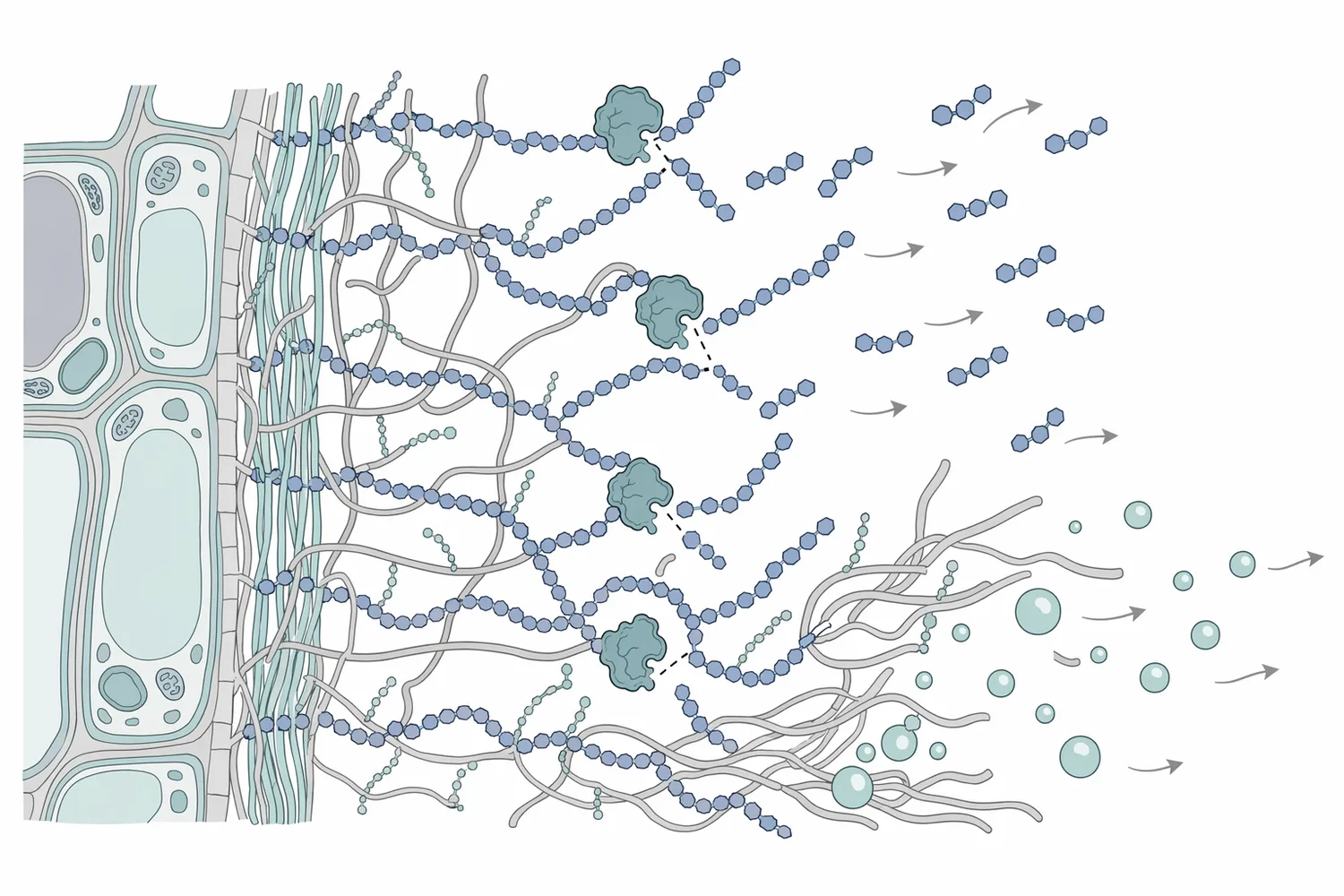
在含 β-葡聚醣比例較高或加工後產生膠體黏度的原料中,β-葡聚醣會增加萃取液黏滯性,使攪拌、泵送、離心、過濾與膜分離負荷提高。穀物產品研究顯示,β-Glucanase 活性會影響 β-葡聚醣分子量降解;分子量下降通常會改變溶液黏度、口感與加工行為,因此同一種酵素作用在不同食品基質中,可能帶來有利或需控制的效果[4]。
對植物萃取產線而言,降黏的價值很實際:較低黏度通常代表固液接觸更均勻、熱與質傳更穩定、過濾壓差較容易控制,也比較不容易形成濾餅堵塞。這不等於所有萃取物都應大幅降解 β-葡聚醣;若產品主打高分子 β-葡聚醣作為膳食纖維或稠度來源,過度水解反而可能削弱標示或質地目的。膳食纖維綜述指出,纖維來源、溶解性、黏度與發酵特性是其營養功能的重要基礎,因此製程中對 β-葡聚醣的處理必須與終端產品定位一致[5]。
β-Glucanase 的核心機制是水解 β-葡聚醣主鏈或支鏈上的 β-糖苷鍵,使原本較長、較能形成黏性網絡的多醣變成較短片段。當聚合度與分子量下降,多醣鏈間纏結減少,溶液黏度、凝膠形成能力與懸浮穩定性可能跟著改變。這類分子量降解在穀物強化 β-葡聚醣產品中已被觀察與討論,顯示酵素活性可直接影響 β-葡聚醣的結構與加工表現[4]。
在細胞壁層級,β-葡聚醣酶的效果可理解為「打開局部通道」。當細胞壁或細胞外基質中的可水解 β-葡聚醣被切短,水合作用與孔隙結構改變,萃取溶劑更容易進入顆粒內部;同時,原本被多醣網絡包埋或與壁材弱結合的多酚、色素、糖苷型風味物質與其他小分子,更容易擴散到液相。木質纖維素水解研究指出,細胞壁結構、木質素特性與酵素吸附會共同影響水解效率,提醒使用者不要把酵素作用簡化為單一受質反應[6]。
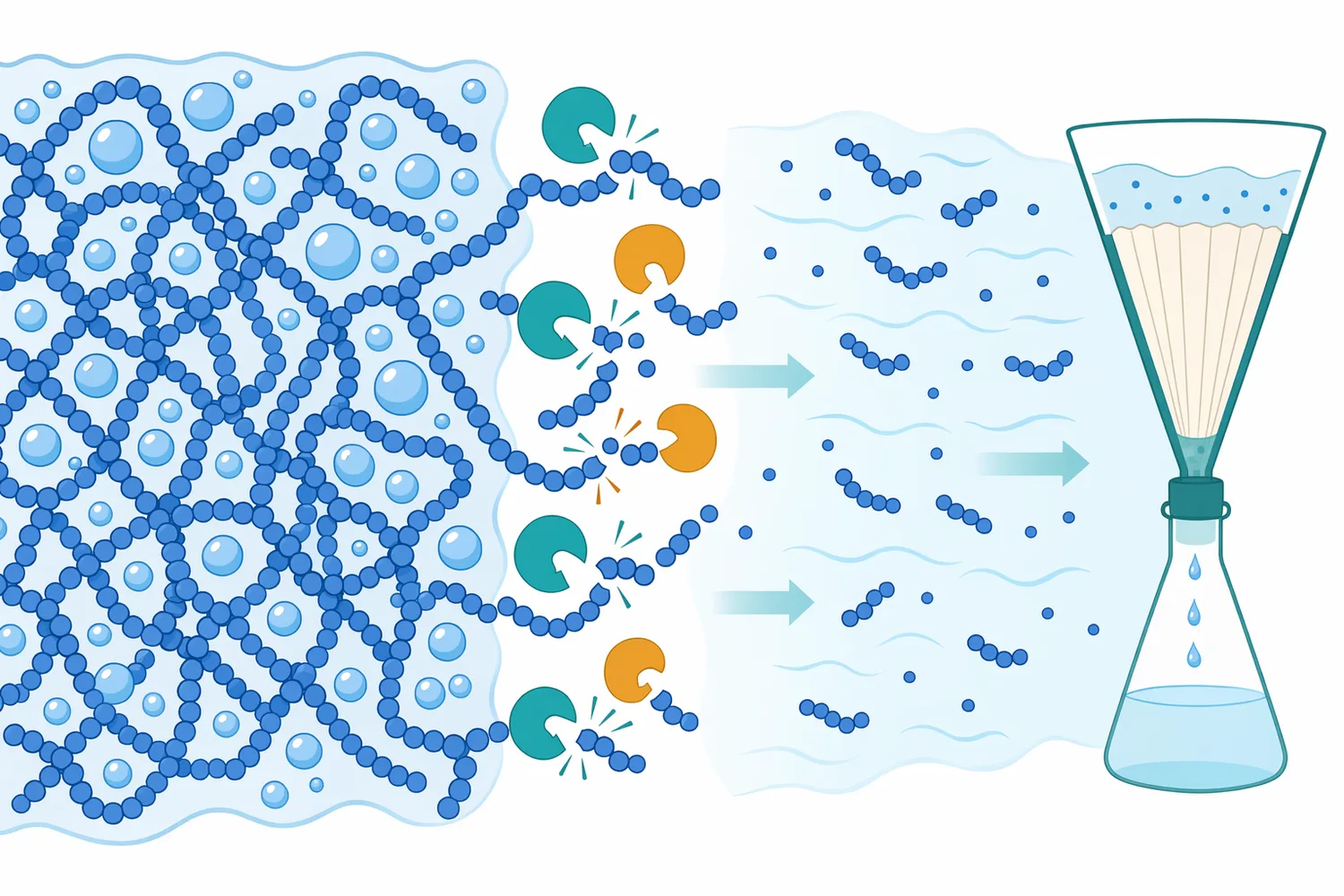
若原料中有酵母、真菌或發酵殘留物,β-Glucanase 還可能作用於微生物細胞壁。酵母與真菌細胞壁常含 β-1,3、β-1,6 葡聚醣等多醣骨架,酵素水解可促進可溶性葡聚醣、細胞壁片段或包埋物釋放;酵素水解酵母葡聚醣以製備水溶性 β-葡聚醣的研究,即顯示水解可改變葡聚醣的溶解性與生物功能表現[7]。
這些機制共同解釋了 β-Glucanase 在植物萃取中的三個主要效益:第一,提升細胞內或細胞壁結合成分的釋放;第二,降低黏度與改善過濾;第三,改變萃取物中多醣分布與澄清行為。不過,這些效益並非固定數值,因為植物基質中限制萃取的關鍵可能是果膠、纖維素、木質素、蛋白質或脂質,而不一定是 β-葡聚醣。
β-Glucanase 較適合用於含有明顯黏稠多醣、穀物 β-葡聚醣、發酵殘留或真菌/酵母細胞壁成分的植物加工系統。例如大麥、燕麥、穀物飲品、植物蛋白副產物流、發酵後植物基料、果渣或草本萃取液,都可能因可溶性多醣造成黏度升高與過濾困難。β-葡聚醣的來源、萃取、純化與工業利用已有完整綜述,顯示其在食品與配料領域兼具機能價值與加工挑戰[1]。
在草本、果蔬與植物多酚萃取中,β-Glucanase 通常不是唯一工具,而是酵素輔助萃取(enzyme-assisted extraction, EAE)的一部分。甜菜根研究將酵素輔助萃取用於總酚、維生素 C 與抗氧化活性評估,說明酵素處理可作為提升植物活性成分回收的可研究路徑;但不同酵素組合、原料粒徑、時間、溫度與溶劑條件都會影響結果[2]。
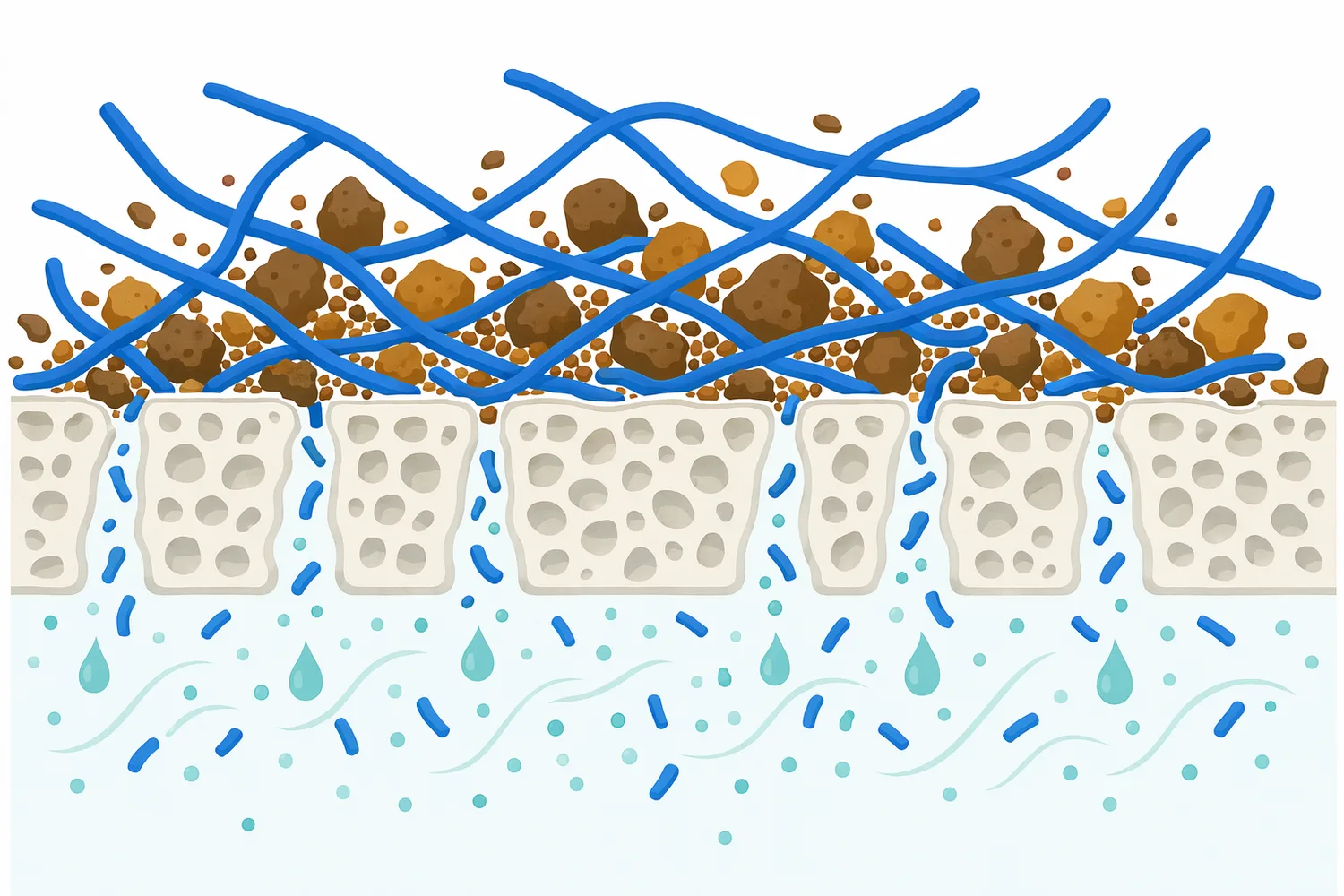
對於富含花青素或天然色素的植物原料,細胞壁鬆解有助於色素釋出,但也可能同時釋放會造成褐變、沉澱或色澤不穩的物質。花青素萃取與表徵綜述指出,食品基質中的花青素受 pH、溫度、溶劑、共色作用與基質交互作用影響;因此在色素萃取中使用 β-Glucanase,應將色澤穩定與澄清需求一併納入流程考量[8]。
在食品副產物再利用方面,如果皮、果渣、葉片、莖部或加工殘渣,β-Glucanase 可協助降低懸浮液黏度,並與熱水、超音波或其他綠色萃取流程搭配。紅枝山茱萸葉與莖的水熱萃取研究顯示,植物副產物流可作為酚類抗氧化物來源,而萃取方式會影響回收效率與濃縮效果[9]。
β-Glucanase 的價值在於溫和、選擇性較高且可嵌入既有濕式製程;但它不是所有萃取問題的通用解方。相較於單純加熱、強溶劑或高能物理破碎,酵素前處理通常更重視基質結構與目標成分穩定性。超音波輔助甜菜根萃取研究顯示,物理強化技術也可促進生物活性成分釋放,代表產業上常見的最佳策略可能是多種溫和技術的組合,而非單一方法取代全部流程[10]。
| 萃取或前處理方式 | 主要作用 | 對植物活性成分的潛在優點 | 主要限制 | 與 β-Glucanase 的關係 |
|---|---|---|---|---|
| β-Glucanase 酵素前處理 | 水解 β-葡聚醣,降低黏度並提高局部通透性 | 有助改善過濾、釋出被多醣網絡包埋的成分 | 對低 β-葡聚醣或高木質化基材效果有限 | 可作為 EAE 的專一性組件 |
| 纖維素酶/半纖維素酶 | 作用於纖維素或半纖維素骨架 | 對細胞壁鬆解更全面 | 可能釋放更多非目標雜質 | 常與 β-Glucanase 互補 |
| 果膠酶 | 降解果膠與中膠層 | 適合果汁、果渣、果皮澄清 | 對穀物 β-葡聚醣降黏不一定足夠 | 可與 β-Glucanase 組合改善澄清 |
| 超音波輔助萃取 | 空化、微射流與顆粒破碎 | 可縮短萃取時間、提高質傳 | 過強條件可能影響熱敏成分 | 可在酵素前後搭配使用 |
| 水熱萃取 | 溫度與水相壓力促進溶出 | 不一定需有機溶劑 | 高溫可能改變色素或香氣 | 酵素可作為較溫和的前段處理 |
酵素與物理技術的搭配需要特別注意順序。若先以高溫或強烈超音波處理,可能改變酵素可作用的受質暴露程度,也可能使酵素失活;若先用酵素軟化細胞壁,再進行溫和物理萃取,則可能提升質傳效率。食品廢棄副產物中酚類回收的超音波輔助與浸泡萃取研究,支持透過溶劑、時間與物理條件調整來改善植物性副產物的酚類回收[11]。
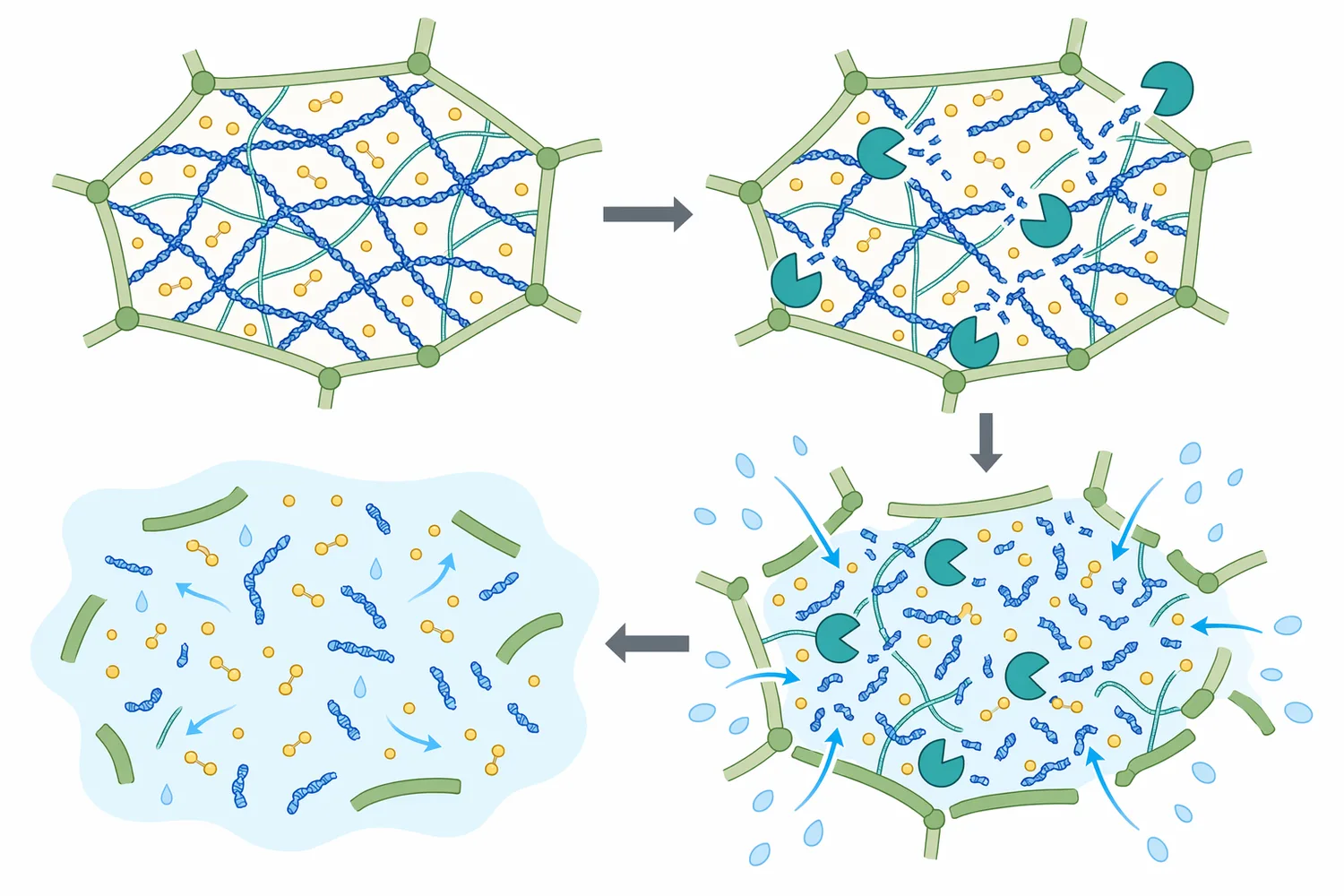
在理想基材中,β-Glucanase 可提升目標成分回收率。這種提升通常來自兩個層面:一是細胞壁局部鬆解使細胞內成分更快進入萃取相;二是高分子膠體降解後,溶液流動性改善,讓混合、熱傳、固液接觸與過濾更穩定。植物細胞壁水解的時間序列觀察指出,水解過程中的形態變化與產率動態密切相關,代表萃取效率往往是「結構變化」與「質傳」共同作用的結果[3]。
在過濾與澄清方面,β-Glucanase 的效益通常最容易被製程端感受到。高 β-葡聚醣或膠體多醣會增加濾液阻力,導致濾速下降、壓差上升或膜面污染加劇;酵素切短多醣鏈後,懸浮液黏度下降,濾餅結構可能較不緊密,後段離心、板框、深層過濾或膜分離的負荷可望降低。β-葡聚醣酶在穀物產品中對分子量降解的研究,提供了這類流變與加工變化的基礎解釋[4]。
不過,提升萃取效率並不永遠等於成品更好。若細胞壁被過度打開,蛋白質、澱粉微粒、核酸、苦味物質、澀味多酚或不穩定色素也可能一起釋出,導致萃取液更混濁、沉澱增加或風味變粗糙。花青素食品基質綜述也提醒,天然色素在不同基質與加工條件下具有穩定性差異,萃取強度過高不一定符合最終品質目標[8]。
因此,β-Glucanase 在 B2B 製程中的定位較像「選擇性鬆解與降黏工具」,而非單純追求最大破壁。對草本萃取、植物飲料、穀物配料、天然色素、果蔬副產物再利用等應用,最佳結果通常是在收率、澄清性、色澤、風味、營養宣稱與後段純化成本之間取得平衡。
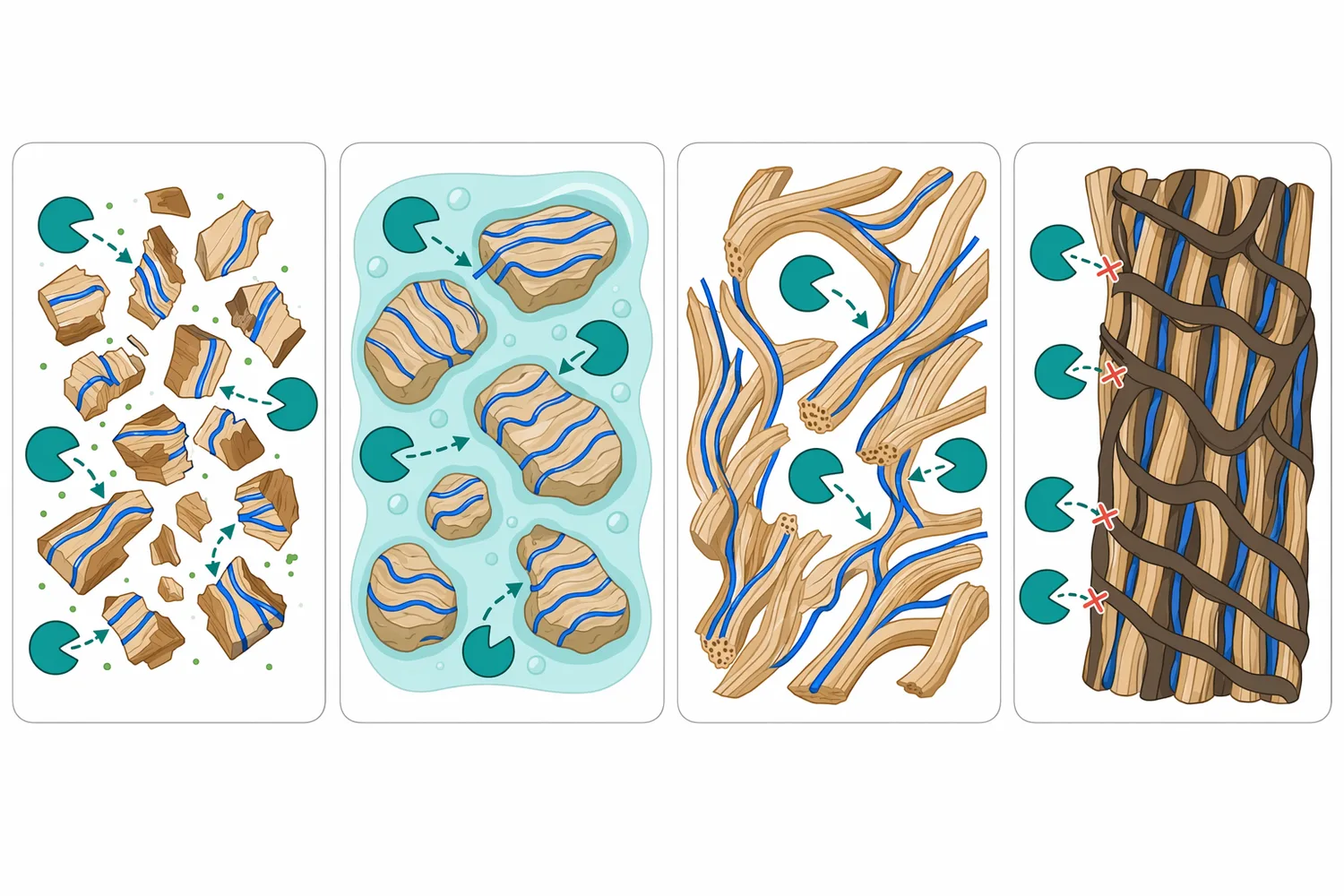
植物細胞壁是複合材料,β-Glucanase 只處理其中一部分結構。若原料限制主要來自纖維素微纖維,纖維素酶可能更關鍵;若來自半纖維素與阿拉伯木聚醣,木聚醣酶更有作用;若是水果或果皮中膠層與果膠造成黏稠,果膠酶通常不可忽略。木質纖維素水解研究顯示,木質素結構會影響纖維素酶吸附與水解效率,說明單一酵素往往會受整體壁材環境限制[6]。
協同的本質在於「先解除一種阻礙,使另一種酵素更容易接近受質」。例如 β-Glucanase 降低可溶性多醣黏度後,攪拌與擴散改善,其他酵素更容易分布到顆粒表面;果膠酶鬆解中膠層後,β-Glucanase 也可能更容易接觸內部多醣。玉米來源 expansin 與酵素水解玉米芯的研究指出,非水解性細胞壁鬆弛因子可促進酵素水解,反映「結構可及性」對酵素效率的重要性[12]。
對食品級植物萃取而言,協同策略應以終端產品需求為核心。若目標是澄清型植物飲料,較強的果膠與 β-葡聚醣降解可能有利;若目標是保留稠度、膳食纖維或口感,則應避免過度降低高分子多醣。若目標是天然多酚或抗氧化萃取物,則需同時觀察有效成分濃度與不欲雜質釋放。植物抗氧化資源的近年綜述也指出,未充分利用的可食植物具有多樣抗氧化潛力,但其萃取與應用仍須依物種與基質特性調整[13]。
在甜菜根、洛神花、莓果、紫色穀物與其他富含色素的植物材料中,萃取效率與色澤穩定同樣重要。甜菜根常被研究作為天然色素與植物活性成分來源,相關研究已探討酵素輔助萃取對總酚、維生素 C 與抗氧化活性的影響;這類案例可作為理解 β-Glucanase 與其他酵素在天然色素原料前處理中角色的參考[2]。
洛神花與其他花青素原料則顯示,萃取方法會影響色素回收、食品添加應用與功能食品潛力。洛神花花青素萃取方法綜述指出,萃取策略需兼顧色素穩定、食品相容性與後續應用;若使用 β-Glucanase,較合理的目的通常是改善基質鬆解與液相流動,而不是直接改變花青素分子本身[14]。
在穀物飲品、燕麥或大麥配料中,β-Glucanase 的角色更接近流變控制。β-葡聚醣是重要的可溶性纖維,但也是造成高黏度、加工困難或口感變化的來源;酵素水解會降低分子量並改變黏度,因此可用於調整泵送性、口感與懸浮穩定。然而,若產品訴求與 β-葡聚醣膳食纖維含量或高分子特性相關,就必須避免不必要的過度降解[5]。
在發酵植物基料、酵母萃取相關副產物或含真菌細胞壁的系統中,β-Glucanase 也可能用於促進細胞壁成分釋放。酵母細胞壁中肝醣與多醣共價連結的研究,顯示微生物細胞壁是結構複雜的多醣網絡;酵素處理可作為解析與改變此類結構的重要工具[15]。
β-Glucanase 通常可安排在萃取前段或萃取初期,讓酵素在水合原料、漿料或低至中等固形物濃度環境中接觸受質。實務上,原料粉碎程度、水分、pH、溫度、攪拌、溶劑比例與停留時間都會影響酵素接觸效率與目標成分穩定性。植物細胞壁酵素水解的 4D 時空資料研究顯示,水解不是瞬間均質反應,而是隨空間位置與時間逐步演變的結構變化[16]。

若流程使用乙醇水溶液、酸化水、熱水或其他食品級萃取介質,需注意酵素對該環境的相容性與目標成分的穩定性。某些多酚或花青素偏好酸性環境,某些酵素則在特定 pH 區間較有效;某些熱水萃取能提高溶出,卻可能使熱敏成分降解。水熱萃取酚類抗氧化物的研究指出,溫度與萃取方式能顯著影響植物材料中酚類濃縮效果,因此酵素條件不能脫離整體萃取策略單獨設定[9]。
在連續或半連續產線中,導入 β-Glucanase 後可能需要重新觀察漿料流動性、濾速、沉降行為與泡沫情形。黏度下降通常有利於泵送與過濾,但顆粒破碎或細胞內容物大量釋出也可能增加微細懸浮物。微流體反應器用於觀察木質纖維素預處理與酵素水解的研究,說明即時觀察結構變化有助於理解前處理與水解之間的關係[17]。
食品級 β-Glucanase 是製程輔助工具,應放在食品安全管理與品質系統中理解,而不是視為終端營養成分。對 B2B 使用者而言,重點在於酵素是否適合既有食品加工條件、是否符合目標市場法規框架,以及是否能在不破壞產品定位的前提下改善製程效率。Enzymes.bio 供應的此產品以 1 kg 單位線上銷售,CoA 與 SDS 隨訂單提供,可作為內部文件建檔、收貨審查與安全資料管理的一部分。
由於 Enzymes.bio 並非製造商或實驗室,使用者不應把供應頁面解讀為完整製程驗證資料;實際導入時仍須依自身原料、設備、配方與地方法規進行判斷。公開文獻能支持 β-Glucanase 在多醣水解、降黏與細胞壁鬆解上的技術原理,但不會自動保證特定植物、特定批次或特定終端產品一定獲得相同結果。
同時,β-Glucanase 的使用也要避免與產品功能訴求衝突。若終端產品強調高黏度口感、完整可溶性纖維或高分子 β-葡聚醣功能,過度水解可能降低產品差異化;若目標是清澈飲料、濃縮萃取物或高效率固液分離,適度水解則可能是有利工具。β-葡聚醣作為可溶性膳食纖維的來源、結構與生物功能差異,正是此類製程決策需要審慎平衡的原因[1]。
β-Glucanase 對富含 β-葡聚醣或相關多醣網絡的基材最有意義;若原料細胞壁主要由果膠、纖維素或高度木質化結構控制,單獨使用 β-Glucanase 的效果可能有限。植物細胞壁水解研究一再顯示,壁材組成、形態與酵素可及性會影響水解結果,因此「同一條件套用所有植物」並不符合科學與產線經驗[3]。
另一個限制是選擇性與雜質釋出的平衡。酵素前處理越強,越可能同時釋出蛋白、多醣片段、微細顆粒、苦澀多酚或色素降解相關物;這些物質可能提高萃取固形物,卻增加澄清、脫色、除味或純化負擔。花青素食品基質研究提醒,天然色素在加工與儲存中受多重因素影響,萃取效率與成品穩定並非同一件事[8]。
最後,若原料或製程涉及發酵副產物,β-Glucanase 作用於酵母或真菌細胞壁可能帶來額外釋出物。這在某些應用中有利,例如提升可溶性細胞壁多醣或風味物質;但在需要清澈、低沉澱或低雜味的產品中,可能需要更謹慎控制處理強度。真菌與植物細胞壁可被酵素複合物水解的研究顯示,不同酵素來源與受質會產生不同程度的細胞壁降解[18]。
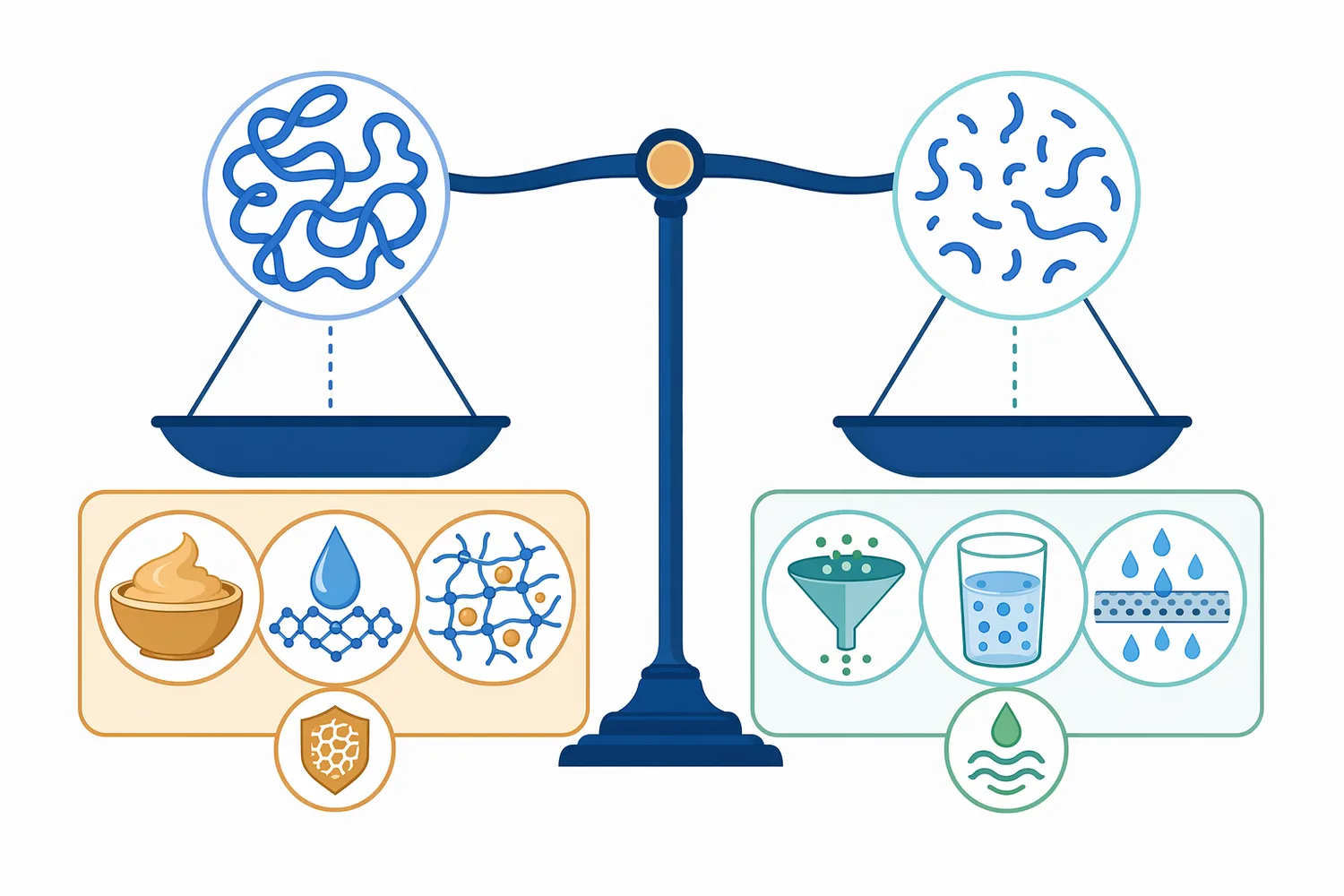
Food Grade β-Glucanase for Plant Extraction 的核心價值在於:以相對溫和的酵素方式切短 β-葡聚醣,降低植物漿料或萃取液黏度,改善溶劑滲透、固液分離與目標成分釋出。對穀物、植物副產物、天然色素、草本萃取、發酵植物基料與含酵母或真菌多醣的系統而言,它可作為酵素輔助萃取中的重要組件,但不應被視為取代所有細胞壁酵素或物理萃取技術的單一解方。
較成熟的導入思路,是先確認製程瓶頸是否與 β-葡聚醣、高黏度膠體或細胞壁通透性有關,再把 β-Glucanase 放入整體萃取策略中,與粒徑、溫度、pH、溶劑、攪拌、過濾及可能的其他酵素協同考量。公開研究支持酵素輔助萃取、超音波、水熱萃取與細胞壁水解等方法在植物活性成分回收上的潛力,但實際效益仍取決於原料與工藝條件[2]。
Enzymes.bio 供應的 Food Grade β-Glucanase for Plant Extraction 以 1 kg 單位線上銷售,並隨訂單提供 CoA 與 SDS;對需要改善植物萃取效率、降低黏度、提升過濾性或優化植物配料製程的 B2B 使用者而言,它是一項可納入製程評估的食品級酵素工具。
以 1 kg 單位販售,現貨供應,可立即出貨。請直接於我們的線上商店下單並付款,我們將為您處理訂單。每筆訂單皆附分析證明書與安全資料表。
購買 Food Grade Β-Glucanase For Plant Extraction →依首次引用順序編號。所有來源皆為開放取用資料,並於發布時確認可連線;正文中的引用編號會連結至此。