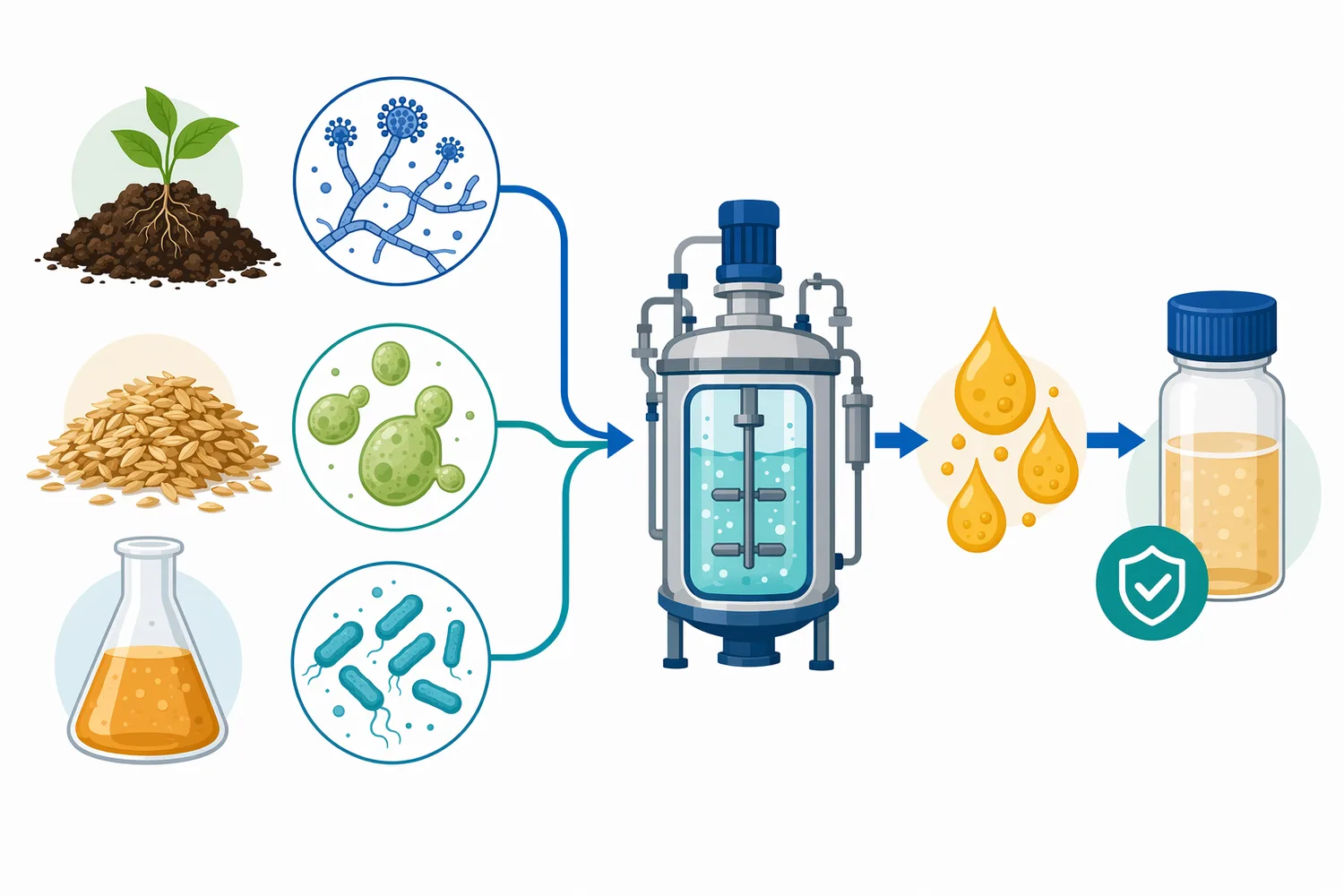
Glucoamylase is an exo-acting starch enzyme that releases glucose from the non-reducing ends of starch-derived chains such as dextrins, maltodextrins, and oligosaccharides. In practical processing, the glucoamylase enzyme is used after starch has been opened up by heat, milling, liquefaction, or alpha-amylase, so that remaining carbohydrate chains can be converted more completely into fermentable glucose. Enzymes.bio supplies glucoamylase directly online by the 1 kg unit; buyers purchase online, the order is processed and shipped, and a Certificate of Analysis and Safety Data Sheet are included with the order.
Glucoamylase, also called amyloglucosidase in many technical contexts, is a carbohydrate-active enzyme used where starch-derived material needs to move closer to glucose. Its main substrate family is not just intact starch, but also the shorter chains produced when starch is cooked, milled, liquefied, or partially hydrolyzed: dextrins, maltodextrins, maltose, maltotriose, and related α-glucans. Reviews of industrial glucoamylase describe it as a key enzyme for starch saccharification because it hydrolyzes α-glucosidic linkages and releases glucose from starch-based substrates [1].
The important word is exo-acting. Alpha-amylase attacks internal α-1,4 bonds within starch chains, rapidly lowering viscosity and creating a mixture of shorter dextrins. Glucoamylase works differently: it acts from the chain end and removes glucose units sequentially. This is why the “glucoamylase vs alpha-amylase” distinction matters in brewing, distilling, bioethanol, and glucose syrup production: alpha-amylase opens the structure and reduces chain length, while glucoamylase continues the conversion toward glucose [1].
Starch itself is a mixture of amylose and amylopectin. Amylose is mostly a linear α-1,4-linked glucose polymer, while amylopectin contains α-1,4 chains with α-1,6 branch points. Those branches slow complete conversion because a linear, end-removing enzyme must work around a branched architecture. Pullulanase and related debranching enzymes are often discussed alongside glucoamylase because they hydrolyze α-1,6 branch linkages, creating more linear chain ends for saccharifying enzymes to attack [2].
At the molecular level, glucoamylase binds the non-reducing end of an α-glucan chain in a catalytic pocket that positions the terminal glucose linkage for hydrolysis. Water participates in cleavage of the glycosidic bond, and the terminal glucose is released while the remaining chain becomes one glucose unit shorter. The process repeats along the chain, which is why glucoamylase is valuable when the desired product is glucose rather than a broad mixture of dextrins [1].
Many fungal glucoamylases include a catalytic domain and, in some forms, a starch-binding region that helps the enzyme associate with insoluble or partially ordered starch surfaces. That structure helps explain why some glucoamylases are more effective on gelatinized starch, while selected “raw starch-digesting” glucoamylases can act more directly on less-pretreated starch granules. Research on a raw-starch-digesting glucoamylase from Penicillium oxalicum specifically evaluated its ability to hydrolyze raw starch and support ethanol fermentation, showing why enzyme structure and substrate accessibility are closely linked in real processes [3].
The mechanism also explains why glucoamylase performance depends strongly on upstream starch preparation. A compact starch granule limits enzyme access; gelatinization, milling, cooking, or liquefaction increases surface area and exposes chain ends. Once alpha-amylase has reduced viscosity and generated soluble dextrins, glucoamylase has more accessible non-reducing ends to process into glucose. This is the practical meaning of the common amylase vs glucoamylase sequence in starch processing: one enzyme rapidly opens the polymer, and the other finishes saccharification toward glucose [1].
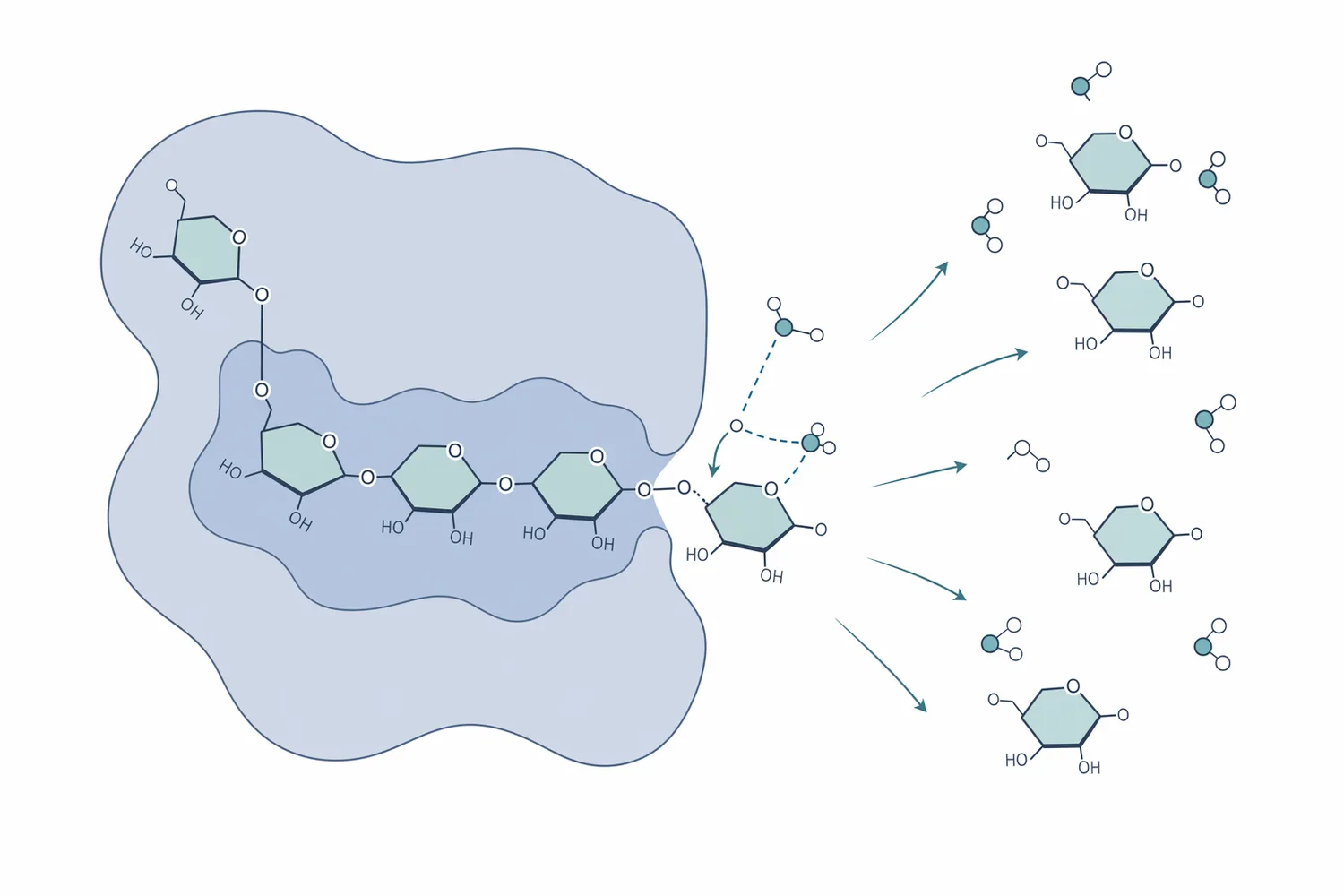
The most useful comparison is not “which enzyme is better,” but which bond and substrate position each enzyme targets. In many starch processes, these enzymes are complementary rather than interchangeable.
| Enzyme | Main action on starch-derived carbohydrates | What changes in the process | Typical role in a starch workflow |
|---|---|---|---|
| Alpha-amylase | Endo-acting cleavage of internal α-1,4 bonds | Long starch chains become shorter dextrins; viscosity drops | Liquefaction and rapid chain-size reduction |
| Glucoamylase | Exo-acting release of glucose from non-reducing chain ends | Dextrins and maltodextrins are converted toward glucose | Saccharification, fermentable sugar generation, glucose syrup production |
| Pullulanase | Debranching cleavage of α-1,6 linkages | Amylopectin branch points are opened, creating more linear chains | Supports more complete hydrolysis when branching limits conversion |
Pullulanase is relevant because amylopectin branching can leave limit dextrins that are less efficiently converted by enzymes acting mainly on α-1,4 chain ends. Reviews of pullulanase describe its role in starch hydrolysis and industrial applications, especially where debranching improves access for saccharification enzymes [2]. In practical terms, glucoamylase can release glucose from available ends, while debranching can create a carbohydrate structure that gives glucoamylase more productive access.
This is also why the search phrases “glucoamylase vs amylase” and “glucoamylase vs alpha amylase” often appear in brewing and distilling discussions. Alpha-amylase does not normally drive a mash all the way to glucose; it produces a dextrin-rich mixture. Glucoamylase is chosen when the objective is higher fermentable sugar generation, lower residual dextrin, or a more complete starch-to-glucose conversion profile [1].
In starch sweetener and fermentation workflows, glucoamylase is generally associated with the saccharification stage. Starch-rich raw materials such as corn, wheat, rice, cassava, potato, sorghum, or other cereal and tuber substrates are first made accessible. Heat and water swell starch granules, mechanical treatment increases surface area, and alpha-amylase may reduce viscosity by breaking large polymers into smaller dextrins. Glucoamylase then converts those dextrins toward glucose [1].
The chemical change is straightforward but commercially important: insoluble or partially soluble starch polymers become soluble, fermentable glucose. Glucose has different functionality from dextrin. It is readily fermented by yeast, contributes sweetness, participates in browning reactions, changes osmotic properties, and can be used as a carbon source for microorganisms in industrial fermentation. This is why glucoamylase appears in applications ranging from brewing and distilling to bioethanol and organic acid production [4].
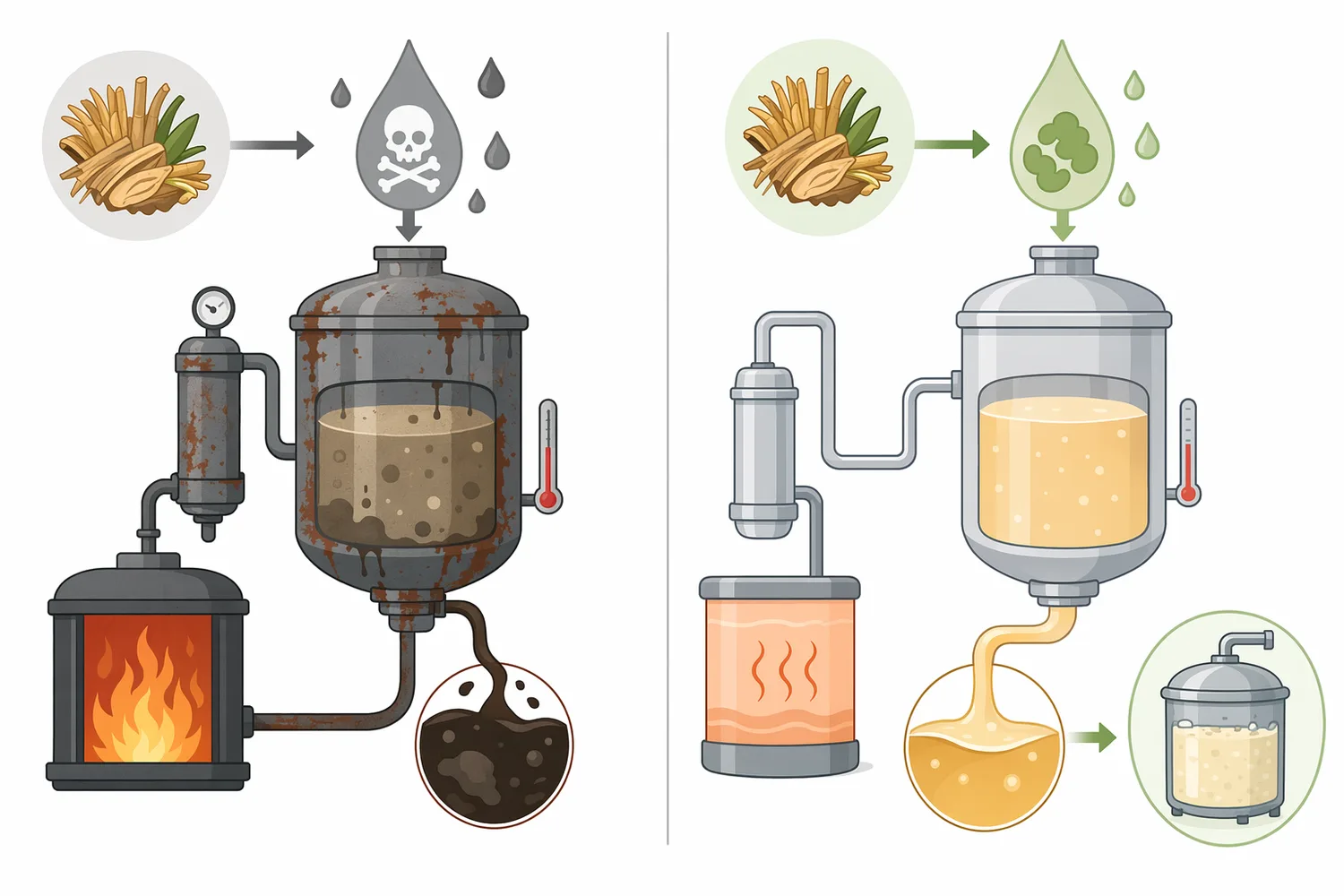
The same mechanism also explains process limitations. If liquefaction leaves inaccessible starch, glucoamylase cannot convert what it cannot physically reach. If pH or temperature conditions are outside the enzyme’s stable operating window, the protein structure can unfold or lose catalytic efficiency. If amylopectin branching is a bottleneck, debranching enzymes may be needed to improve access. Glucoamylase is therefore best understood as a targeted saccharification enzyme, not a universal replacement for starch preparation [2].
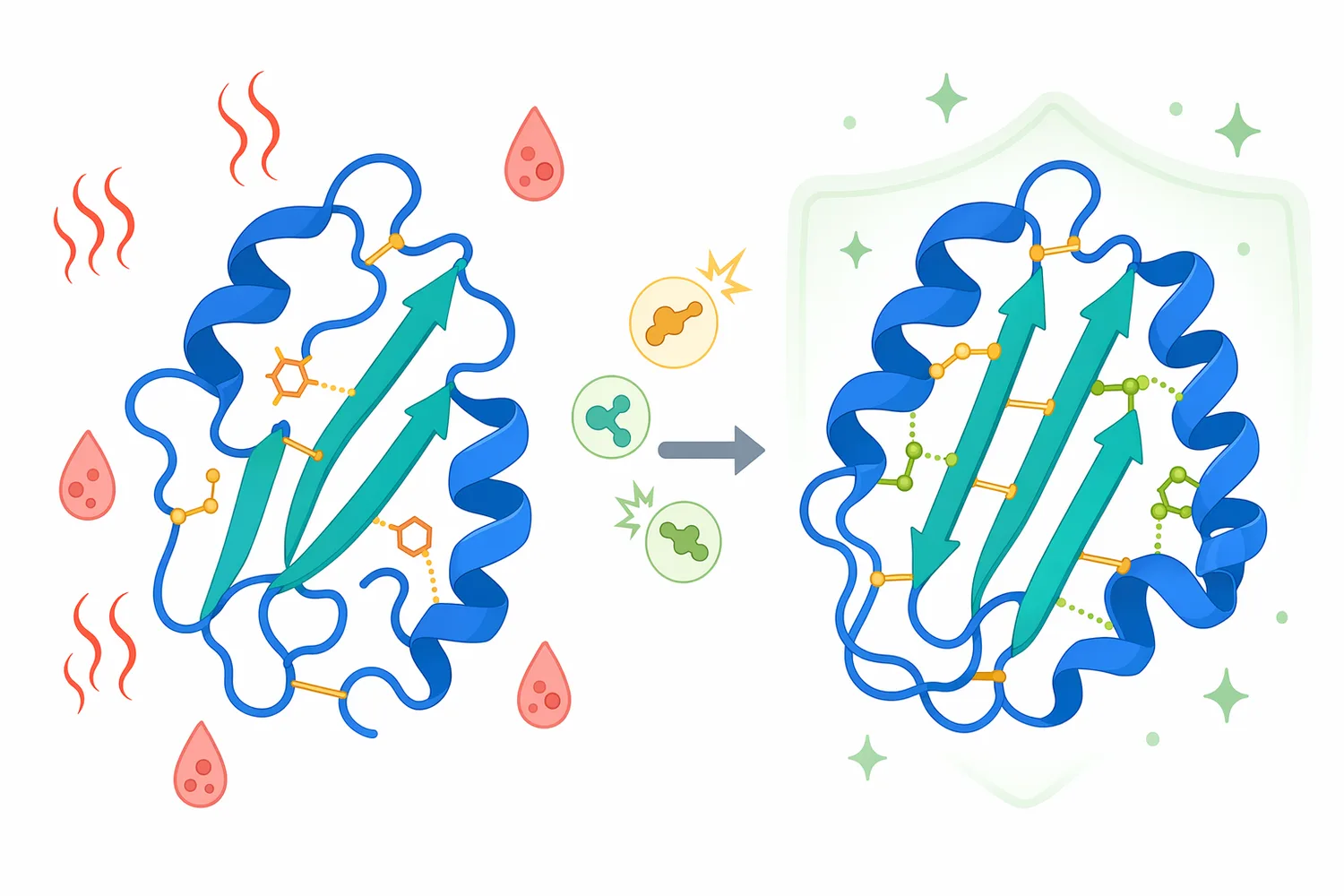
Searches for “glucoamylase temperature range” usually come from users trying to understand whether the enzyme will fit a warm mash, saccharification tank, distilling process, or food formulation. Published glucoamylases are commonly investigated under acidic to mildly acidic conditions and warm processing temperatures, but the exact useful range depends on the microbial source, enzyme structure, formulation, substrate, and process design. A 2021 study on Talaromyces leycettanus glucoamylase used site-directed mutagenesis specifically to improve thermostability and catalytic efficiency for industrial saccharification, illustrating how temperature tolerance is a major research target for this enzyme class [5].
Thermostability matters because starch processes often operate warm enough to improve solubility and reduce contamination risk, but proteins can denature if the temperature exceeds what the enzyme structure can tolerate. When glucoamylase unfolds, the active site no longer positions the glycosidic bond correctly, and glucose release slows or stops. Protein-engineering research focuses on strengthening structural interactions—such as salt bridges, hydrogen-bond networks, or local packing—so the enzyme retains its active conformation for longer under saccharification conditions [6].
Acid tolerance is also valuable. Many fungal glucoamylases are naturally suited to acidic environments, which can match conditions used in starch saccharification, beverage production, and some industrial fermentations. An acid-thermostable glucoamylase from Aspergillus japonicus was characterized for potential use in paper bio-deinking, showing that glucoamylase research extends beyond food and fuel into industrial processes where low pH and heat stability can be beneficial [7].
For distilling, the phrase “glucoamylase enzyme distilling” usually refers to using glucoamylase to increase the fermentable glucose available from grain starch. Grain mashes contain starch that must be converted into sugars before yeast can produce ethanol efficiently. Alpha-amylase can reduce viscosity and create dextrins; glucoamylase then releases glucose from those dextrins so yeast has a more directly fermentable carbon source [1].
In high-attenuation brewing, glucoamylase can reduce residual dextrins that would otherwise remain after mash enzymes and yeast fermentation. The practical effect is a drier, more fully attenuated carbohydrate profile when the process is designed for that outcome. This does not mean every beer should use glucoamylase; residual dextrin can be desirable for body and mouthfeel. The enzyme is most relevant when the product target favors lower residual carbohydrate and greater fermentability [1].
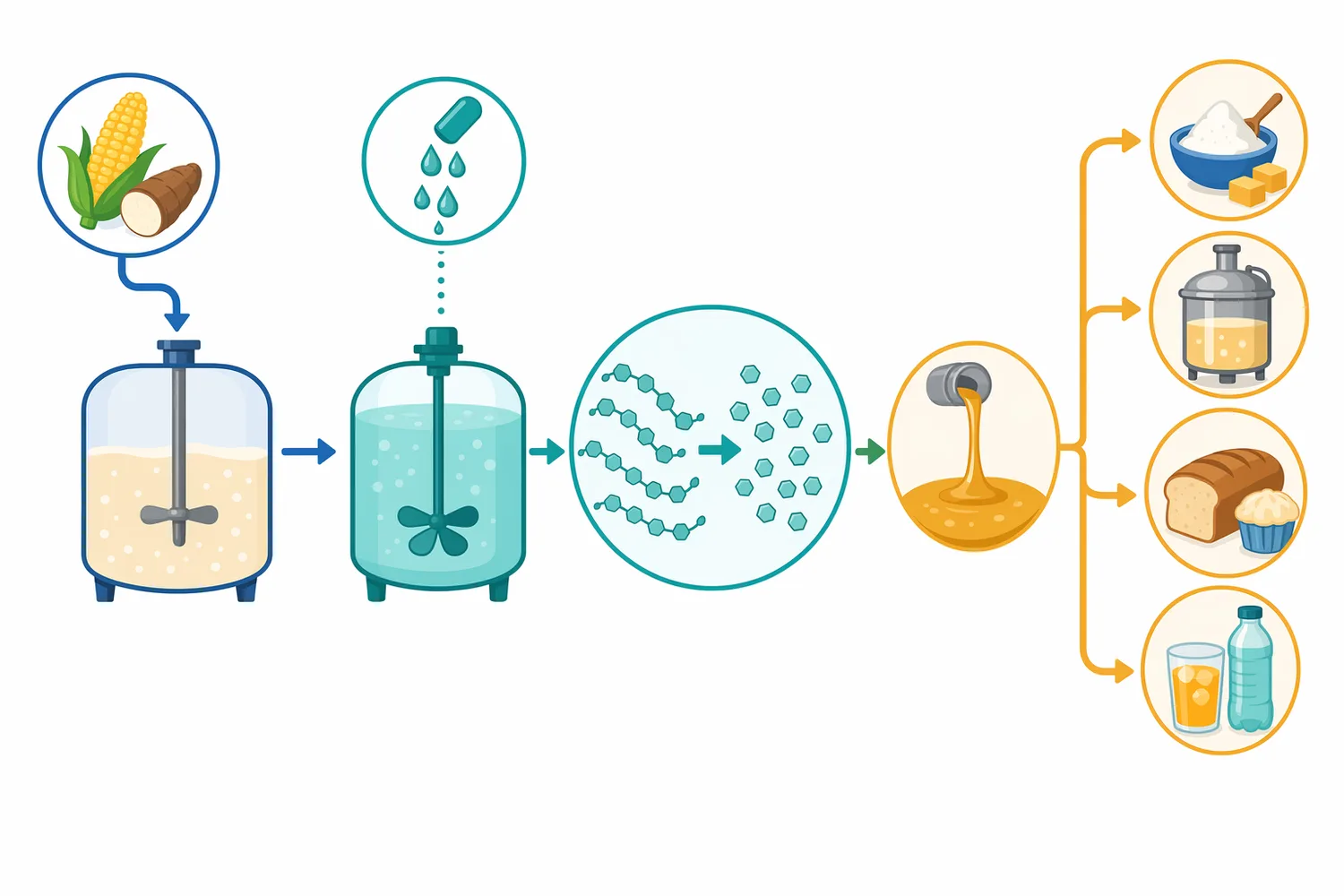
In distilling, residual dextrin is usually less valuable than fermentable sugar because the goal is ethanol production rather than finished beverage body. Research on raw-starch-digesting glucoamylase from Penicillium oxalicum connected raw starch hydrolysis with ethanol fermentation, demonstrating the industrial interest in enzymes that can convert starch-rich substrates into fermentable sugars with fewer or more efficient pretreatment steps [3].
Bioethanol production depends on converting starch-rich biomass into fermentable sugars. Glucoamylase contributes by transforming liquefied starch fractions into glucose, which yeast and other microorganisms can metabolize into ethanol or other fermentation products. The enzyme’s business value in this setting is not simply “more enzyme,” but more complete conversion of prepared starch into a sugar profile that the fermentation organism can use [3].
Industrial fermentation also includes products beyond ethanol. Enzyme-assisted conversion of agro-industrial residues is discussed in the context of itaconic acid production, where biomass-derived carbohydrates must be made available as microbial carbon sources. Glucoamylase is relevant in this broader bioprocessing logic because starch-rich residues or cereal by-products often require enzymatic hydrolysis before fermentation organisms can efficiently consume them [4].
The substrate determines how much help glucoamylase can provide. A clean liquefied starch stream gives the enzyme a different environment from whole grain mash, bran-rich slurry, or mixed agro-industrial residue. Fibers, proteins, lipids, minerals, and particle size can all influence water availability and enzyme access. This is why research frequently studies glucoamylase alongside production organisms, pretreatment methods, and substrate types rather than as an isolated ingredient [8].
In food processing, glucoamylase is used where controlled glucose generation from starch is beneficial. In glucose syrup production, the purpose is direct: convert liquefied starch into a glucose-rich syrup. In cereal processing and baking, the role is more nuanced. Glucose can feed yeast, contribute to crust browning through Maillard reactions, influence sweetness, and change the balance between fermentable and non-fermentable carbohydrates [1].
In baking, too little starch hydrolysis may limit yeast nutrition, while too much can affect dough handling, stickiness, or texture. Glucoamylase therefore functions as a precision tool for carbohydrate profile adjustment rather than a generic “improver.” It acts on starch-derived chains that are available in the dough system, releasing glucose over time and changing what yeast and browning chemistry can use [8].

Solid-state fermentation research with Aspergillus oryzae cultivated on wheat bran shows why cereal substrates are important in enzyme production and application science. Wheat bran is rich in complex plant material, and studies of fungal enzyme production on such substrates often include glucoamylase, alpha-amylase, and cellulase because cereal matrices contain starch, fiber, and associated nutrients that interact during processing [8].
Not all glucoamylases behave the same on raw starch. Gelatinized starch is swollen and more accessible, while raw starch granules have semi-crystalline regions that resist enzymatic attack. A raw-starch-digesting glucoamylase must not only catalyze glycosidic bond hydrolysis; it must also bind effectively to a granular substrate surface and act at accessible points without relying entirely on prior gelatinization [3].
The Penicillium oxalicum raw-starch-digesting glucoamylase study is important because it connects enzyme characteristics with ethanol fermentation, a practical end use where heating energy, substrate handling, and conversion efficiency matter. When an enzyme can hydrolyze raw or less extensively treated starch, it may support process designs that differ from conventional high-temperature gelatinization and liquefaction routes [3].
However, raw starch digestion should not be generalized across all glucoamylases. The presence or absence of starch-binding regions, the condition of the raw material, particle size, botanical source, moisture level, and process temperature all affect performance. For most industrial starch conversion, glucoamylase is still most effective when the substrate has been made accessible by cooking, milling, liquefaction, or other pretreatment [1].
Commercial and research glucoamylases are commonly associated with fungi such as Aspergillus, Rhizopus, Mucor, Talaromyces, and Penicillium, although bacterial glucoamylases are also studied. Research on Aspergillus niger glucoamylase expression in Pichia pastoris reflects a broader trend: known glucoamylase genes can be cloned and expressed in production hosts to study enzyme properties and improve output [9].
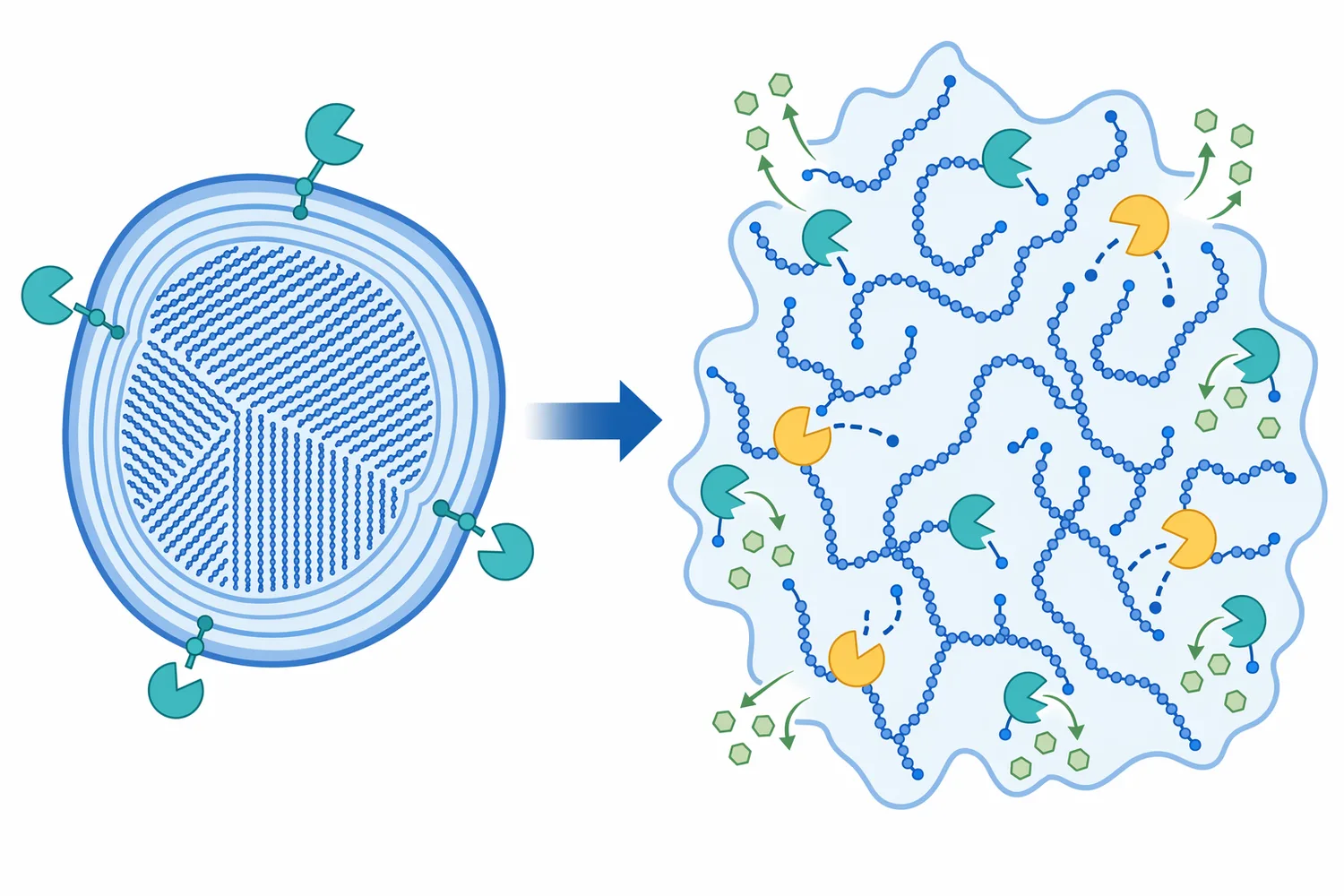
Aspergillus niger is especially prominent in industrial enzyme literature. A study on overexpressing NADH kinase in A. niger examined effects on glucoamylase production, showing that cellular metabolism and redox balance can influence enzyme output in fungal systems [10]. This type of work matters because glucoamylase availability at industrial scale depends not only on enzyme chemistry but also on biological production efficiency.
Other organisms are being explored for different performance profiles. Studies have reported extracellular glucoamylase from Paenibacillus amylolyticus and glucoamylase production by Mucor indicus, Mucor hiemalis, and Rhizopus oryzae under solid-state fermentation. This diversity matters because microbial source can influence pH behavior, temperature tolerance, substrate preference, stability, and suitability for different applications [11].
Fruit-waste soil and agro-industrial residues have also been used as screening and production contexts for glucoamylase-producing microorganisms. A 2025 study isolated and optimized glucoamylase-producing bacteria from fruit-waste soil for industrial applications, reinforcing the ongoing search for enzymes adapted to different feedstocks and operating environments [12].
Because starch saccharification can involve heat, acidity, long residence times, and high solids, enzyme stability is a major development target. Protein engineering can alter amino acid residues to improve thermostability, catalytic efficiency, or resistance to process stress. The Talaromyces leycettanus glucoamylase mutagenesis study is an example of this approach, focusing on improved thermostability and catalytic efficiency for industrial saccharification [5].
Protein-engineering reviews emphasize that industrial enzymes are often improved by changing structure-function relationships rather than simply discovering new natural enzymes. For glucoamylase, that may mean stabilizing the catalytic domain, improving substrate binding, or reducing loss of activity under warm acidic conditions. Mechanistically, these changes aim to keep the enzyme folded correctly and maintain the active-site geometry needed to hydrolyze glycosidic bonds [6].
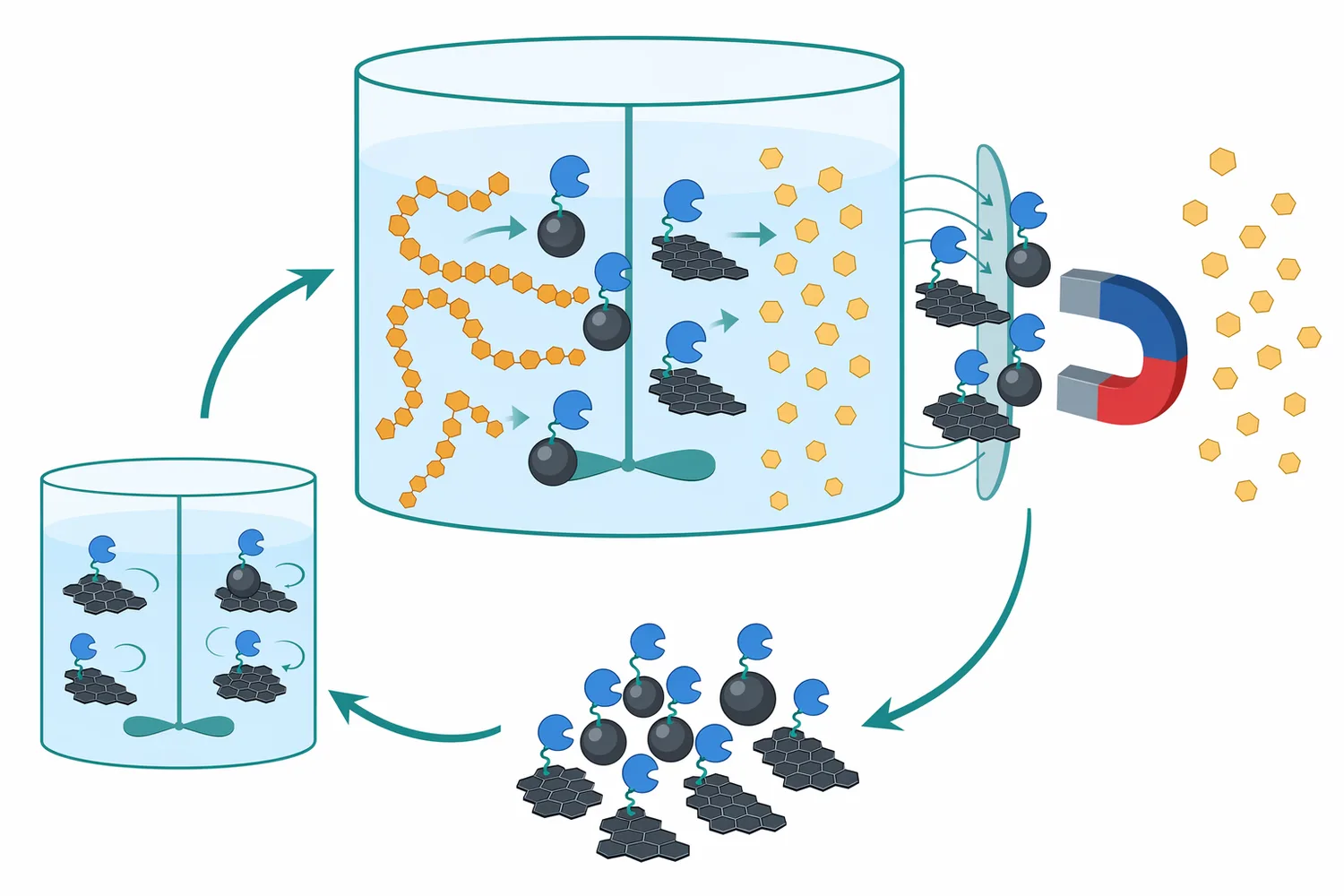
Immobilization is another research route. Immobilized glucoamylase can be attached to a solid support so that it is easier to separate from the reaction medium or reuse in repeated cycles. A study immobilized glucoamylase on a triazine-functionalized magnetic graphene oxide composite and reported improved stability and reusability, showing how materials engineering can change enzyme handling and process durability [13].
Searches such as “what is glucoamylase,” “what does glucoamylase do,” and “glucoamylase supplement” often come from nutrition and digestive enzyme contexts rather than industrial processing. In that setting, glucoamylase is usually described as supporting the breakdown of starch-derived carbohydrates into glucose. The mechanism is the same as in food and fermentation systems: the enzyme hydrolyzes α-glucan chain ends to release glucose [1].
It is important to distinguish industrial glucoamylase from maltase-glucoamylase, sometimes written “maltase glucoamylase,” which refers to a human intestinal enzyme complex involved in digestion of dietary starch products. Searches for “glucoamylase deficiency” or “glucoamylase low” generally relate to human digestive physiology or clinical interpretation, not to how a processing enzyme performs in a mash, syrup, feed, or food application. Enzymes.bio’s glucoamylase should therefore be understood as an enzyme ingredient for starch breakdown, not as medical advice or a disease-related product.
For supplement or nutrition-product applications, claims must be handled carefully. It is scientifically reasonable to say that the enzyme glucoamylase helps break starch-derived carbohydrates into glucose. It is not responsible to claim that glucoamylase alone treats digestive disease, corrects a deficiency, or guarantees a specific health outcome. The strongest, most transferable claim remains the biochemical one: glucoamylase supports starch and dextrin hydrolysis toward glucose [1].
Although glucoamylase is most strongly associated with starch sweeteners and fermentation, it also appears in other industrial applications where starch or starch-derived residues are present. In paper bio-deinking, starch-based coatings, sizing materials, or contaminants can interfere with processing. An acid-thermostable glucoamylase from Aspergillus japonicus was evaluated for paper bio-deinking potential, showing how starch-degrading enzymes can contribute outside conventional food and fuel pathways [7].
In feed processing, glucoamylase may be used where starch-rich ingredients need greater enzymatic breakdown. The practical effect depends on animal species, feed matrix, heat treatment, moisture, retention time, and the presence of other enzymes. The mechanism is still glucose release from starch-derived chains, but the performance outcome is tied to the total feed formulation and processing environment rather than the enzyme alone [1].
Agro-industrial residues are another area of interest. Enzyme-driven processing can help convert starchy by-products into fermentable sugars or microbial substrates, reducing waste while creating value. Research on enzyme-assisted use of agro-industrial waste in itaconic acid production illustrates the broader industrial theme: enzymes such as glucoamylase help transform complex carbohydrate residues into more usable carbon sources [4].
The first practical benefit is more complete saccharification. When starch has already been opened into dextrins, glucoamylase can continue hydrolysis toward glucose rather than leaving a dextrin-rich carbohydrate profile. This matters for glucose syrup, brewing attenuation, distilling yields, and fermentations where glucose is a preferred carbon source [1].
The second benefit is improved fermentable sugar availability. Yeast and many industrial microorganisms readily use glucose, while longer dextrins may remain partly unfermented depending on the organism. In distilling and ethanol applications, converting dextrins into glucose can improve the amount of carbohydrate that is available for fermentation, provided the rest of the process supports microbial performance [3].
The third benefit is carbohydrate profile control. In some products, residual dextrin provides body, viscosity, or mouthfeel; in others, it is an inefficiency. Glucoamylase helps shift the balance toward glucose when that is the desired process target. This is why it can be valuable in one brewing application and unnecessary or excessive in another [1].
The fourth benefit is compatibility with multi-enzyme systems. Starch conversion rarely depends on a single enzyme alone. Alpha-amylase, glucoamylase, pullulanase, cellulase, protease, and other enzymes may each address different components of a raw material. Studies of fungal cultivation on wheat bran, for example, commonly evaluate multiple enzymes because cereal substrates contain starch plus non-starch polysaccharides and proteins [8].
Glucoamylase can be expected to hydrolyze accessible starch-derived α-glucans toward glucose under suitable conditions. It is especially useful after liquefaction, cooking, milling, or other treatment that exposes chain ends and makes substrate accessible. Its core value is specific, concrete, and measurable in process terms: more glucose from dextrins and related starch fragments [1].
Glucoamylase should not be expected to solve every starch-processing limitation by itself. It does not physically mill grain, gelatinize starch granules, remove all branch-point limitations, or guarantee yeast performance. If the substrate is poorly hydrated, highly crystalline, insufficiently liquefied, or constrained by non-starch matrix components, the enzyme’s access to glycosidic bonds may be limited [3].
It should also not be treated as interchangeable with alpha-amylase. The common phrase “glucoamylase vs alpha-amylase” exists because the enzymes solve different parts of the starch conversion problem. Alpha-amylase rapidly reduces chain length and viscosity; glucoamylase releases glucose from chain ends; pullulanase opens α-1,6 branch points. A complete starch strategy often uses these functions in sequence or combination [2].
Enzymes.bio supplies glucoamylase directly online by the 1 kg unit. The buying process is simple: the product is purchased online, the order is processed, and the enzyme is shipped. A Certificate of Analysis and Safety Data Sheet are included with the order.
For users working with starch conversion, fermentation, distilling, food processing, feed, or nutrition-oriented formulations, glucoamylase is best understood as a practical enzyme for converting accessible starch-derived chains into glucose. Its value comes from a well-established mechanism: endwise hydrolysis of dextrins and related α-glucans, producing glucose that can be fermented, formulated, or further processed [1].
The key takeaway is straightforward: glucoamylase does not merely “break down starch” in a generic way. It completes the conversion work left after starch has been opened up, removing glucose units from chain ends and reducing residual dextrin when the process target calls for higher glucose formation. That specific role is why glucoamylase remains central to saccharification, brewing, distilling, bioethanol, glucose syrup, cereal processing, and starch-focused enzyme blends.
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Glucoamylase →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.