マンナナーゼ酵素は、飼料中のβ-マンナン、ガラクトマンナン、グルコマンナンなどのマンナン系多糖を加水分解し、植物性原料の栄養利用性と腸内環境を支援する動物飼料添加物です。特にブロイラー、採卵鶏、離乳子豚、育成・肥育豚などの単胃動物では、β-マンナンが消化管粘性、未消化栄養素、飼料誘導性免疫応答に関与し得るため、β-マンナナーゼの利用意義が明確です[1]。Enzymes.bioは本品を1kg単位でオンライン供給するサプライヤーであり、注文時にCoAおよびSDSが併せて提供されます。
飼料用マンナナーゼ、特にβ-マンナナーゼは、植物細胞壁や種子貯蔵多糖に含まれるβ-1,4-マンナン骨格を切断する外因性酵素です。トウモロコシ・大豆粕主体の飼料であっても、大豆粕、パーム核粕、コプラミール、グアー関連原料などにはマンナン系非デンプン性多糖が含まれ、単胃動物の内因性消化酵素だけでは十分に分解されません。飼料用外因性酵素は、動物側の消化能力では処理しにくい基質に作用し、栄養素の放出、消化管内容物の性状改善、腸内環境への二次的影響を目的に使われます[2]。
β-マンナンは単なる「繊維」ではなく、飼料設計上は抗栄養因子として扱うべき成分です。可溶性画分は消化管内容物の粘性を高め、消化酵素と基質の接触、栄養素の拡散、吸収面への移動を妨げる可能性があります。不溶性画分も植物細胞壁の物理的障壁として働き、タンパク質、脂質、デンプンなどへのアクセスを制限します。離乳子豚を対象とした研究では、β-マンナナーゼが腸管健康と成長に関連する栄養的・機能的役割を持つことが検討されており、マンナン分解を通じた消化管負荷の低減が主要な論点になっています[1]。
飼料中のマンナン量が増える場面では、マンナナーゼの実務的価値が高まります。たとえば、原料価格や調達事情により副産物原料の比率を高める場合、パーム核粕やコプラミールのようなマンナン系多糖を多く含む原料を使う場合、または大豆粕比率が高い若齢動物用飼料を設計する場合です。コプラミールをマンナナーゼと酵母で栄養改善する研究では、難消化性マンナンを含む原料の利用性を高める方向性が示されており、マンナナーゼは副産物原料の栄養価を引き出す技術として理解できます[3]。
β-マンナン、とくにガラクトマンナンや一部の水溶性マンナンは、水を抱え込んで消化管内容物の粘性を高めます。粘性が上がると、消化酵素がデンプン粒、タンパク質、脂質に到達しにくくなり、胆汁酸やリパーゼの拡散、単糖・アミノ酸・脂肪酸の吸収面への移動も遅くなります。その結果、消化管後部へ流れる未消化栄養素が増え、微生物発酵の基質が過剰になりやすくなります。ブロイラーを対象としたβ-マンナナーゼのメタ解析では、エネルギーおよび栄養素利用への影響が検討されており、マンナン分解が飼料エネルギー評価と関係することが示されています[4]。
粘性の問題は、単に栄養吸収効率だけでなく、敷料状態、糞便性状、腸内微生物の基質供給にも波及します。未消化の炭水化物やタンパク質が後腸に流入すると、乳酸産生菌や短鎖脂肪酸産生菌に利用される場合もありますが、条件によっては望ましくない発酵、アンモニア生成、腸管刺激につながります。外因性酵素のレビューでは、酵素添加物が栄養消化と腸内環境の両面に関与することが整理されており、マンナナーゼは標的基質が明確な炭水化物分解酵素として位置づけられます[2]。
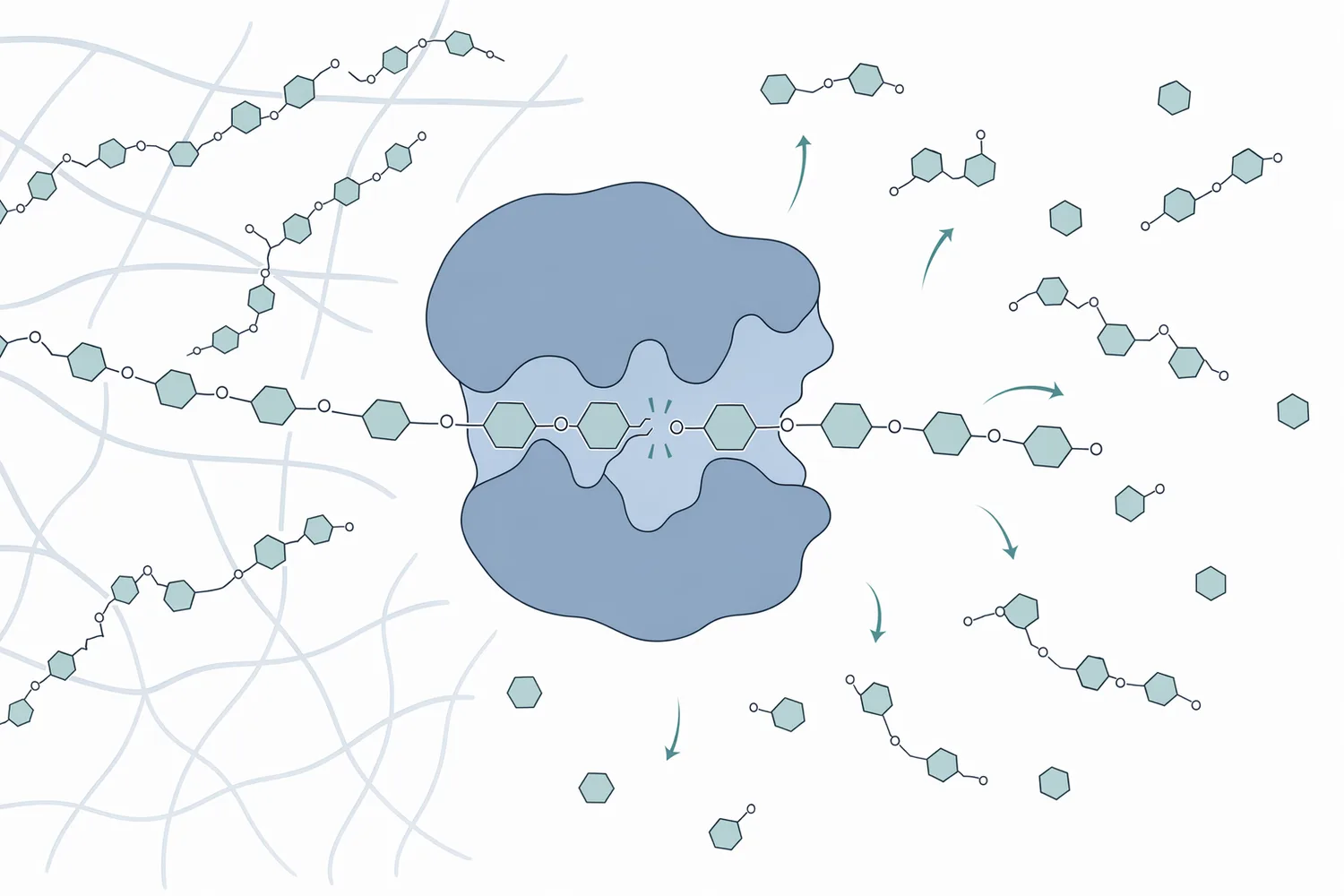
β-マンナンの重要な特徴は、栄養学的な粘性だけでなく、免疫系への影響が議論されている点です。マンナン様構造は微生物細胞壁にも存在するため、腸管免疫系がこれを「微生物関連分子パターン」に近いシグナルとして認識する可能性があります。この反応が過剰または持続的に起こると、栄養素が成長や産卵ではなく免疫応答、急性期反応、上皮修復へ振り向けられる可能性があります。離乳子豚の腸管健康に関するβ-マンナナーゼ研究では、マンナン分解が成長性能だけでなく腸内環境・免疫関連指標と結びつけて検討されています[1]。
この文脈で、β-マンナナーゼは「免疫を刺激する添加物」ではなく、未分解β-マンナンによる不要な刺激を減らす方向に働く酵素として説明するのが適切です。大きなマンナンポリマーを短いマンノオリゴ糖へ切断することで、粘性を下げ、細胞壁構造を緩め、免疫受容体に提示される高分子構造を変化させる可能性があります。育成豚のトウモロコシ・大豆粕飼料でβ-マンナナーゼ単独またはマルチカルボヒドラーゼ複合体との併用を検討した研究では、栄養代謝と腸管健康の双方が評価対象となっており、単なる繊維分解以上の作用が注目されています[5]。
マンナナーゼがβ-マンナンを分解すると、マンノオリゴ糖などの低分子画分が生じます。これらは動物自身の消化吸収に直接使われるだけでなく、腸内細菌の発酵基質として働く可能性があります。短鎖脂肪酸産生、乳酸菌やその他の有用菌の相対的増加、病原性細菌に利用されやすい未消化栄養素の減少などは、マンナナーゼの二次的な腸内環境支援として説明されます。ただし、腸内微生物叢は飼料組成、日齢、衛生状態、抗菌剤使用歴、疾病圧に強く左右されるため、一定の菌叢変化を保証するものではありません[5]。
ブロイラー腸内の食品媒介病原体定着を抑える栄養介入に関するシステマティックレビューでは、飼料添加物や栄養設計が腸内微生物を介して衛生リスクに影響し得ることが整理されています。マンナナーゼは直接的な殺菌剤ではありませんが、未消化基質の流入や腸管内容物の物性を変えることで、病原体が増殖しやすい環境を間接的に変える可能性があります[6]。
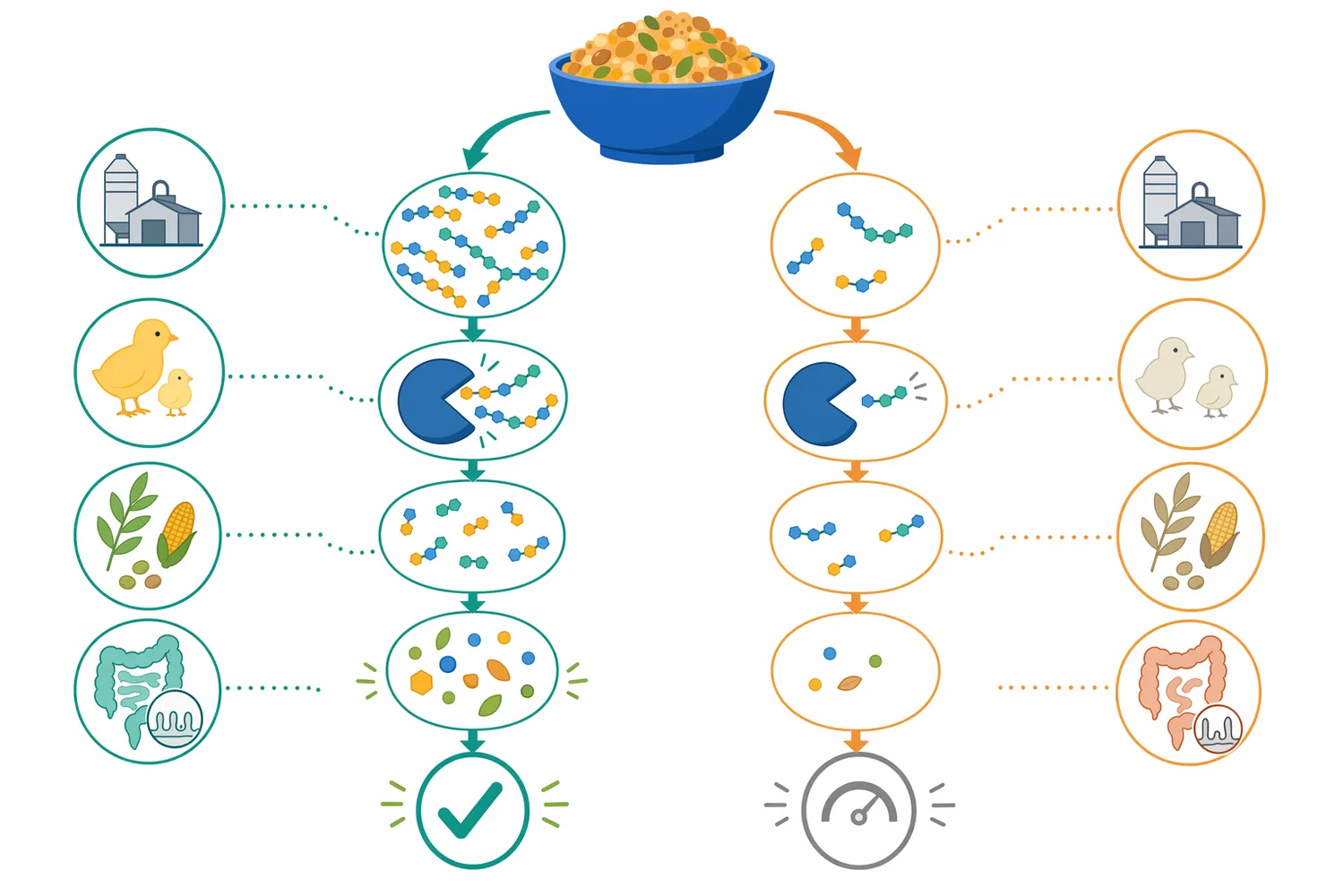
β-マンナナーゼの一次作用は、マンナン主鎖のβ-1,4-グリコシド結合を加水分解することです。エンド型に作用するβ-マンナナーゼは、高分子マンナンの内部結合を切断し、分子量を下げ、マンノオリゴ糖や短鎖多糖へ変換します。この段階で粘度低下、細胞壁マトリクスの緩み、他の消化酵素がアクセスできる表面積の増加が起こります。外因性酵素の飼料利用に関するレビューでは、酵素の効果は標的基質と作用部位の一致に依存すると説明されており、マンナナーゼではβ-マンナンの存在が効果発現の前提になります[2]。
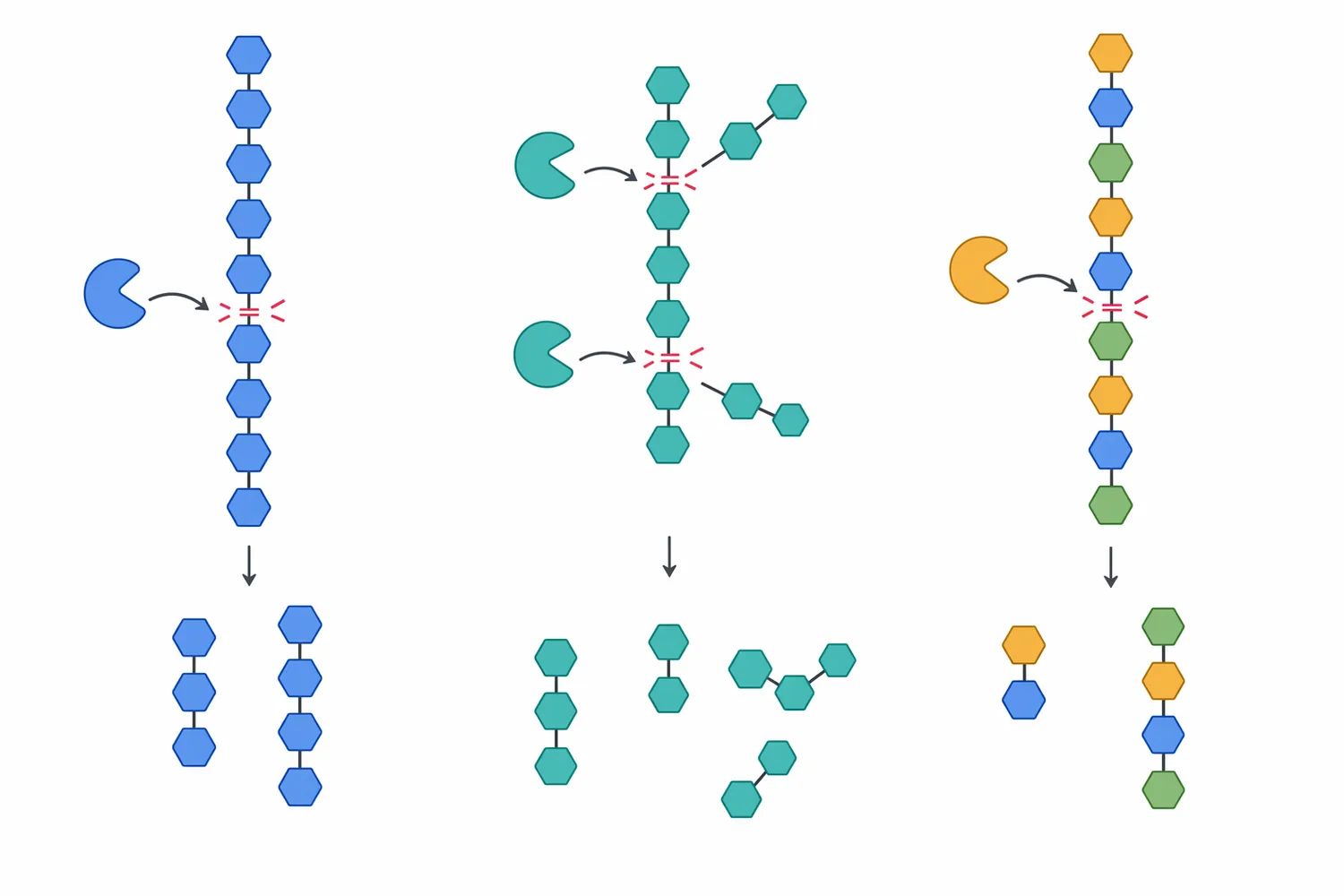
次に、飼料マトリクスの物理構造が変化します。植物細胞壁はセルロース、ヘミセルロース、ペクチン、リグニン、タンパク質、多糖複合体からなるため、マンナン部分の分解だけでも細胞壁の密度や水保持性が変わります。これにより、膵液由来アミラーゼ、プロテアーゼ、リパーゼ、胆汁酸が基質に届きやすくなり、栄養素の見かけの消化率や代謝エネルギー利用に影響する可能性があります。ブロイラーにおけるβ-マンナナーゼ補給のメタ解析がエネルギー・栄養素利用を対象としているのは、この物理化学的作用が成績指標へ反映され得るためです[4]。
さらに、腸管上皮と免疫系への負荷が変わります。高粘性内容物や未消化栄養素の増加は、腸管上皮の酸化ストレス、粘膜刺激、微生物由来代謝産物の変動を通じて、絨毛構造やバリア機能に影響し得ます。β-マンナナーゼがマンナンを低分子化すれば、腸管表面に接触する高分子抗栄養因子の量が下がり、上皮細胞のエネルギー配分や炎症関連反応が変化する可能性があります。全肥育期間の豚を対象としたβ-マンナナーゼ添加研究では、生産成績と腸管健康が同時に評価されており、消化性と腸内状態の接点が示されています[7]。
ブロイラーでは、飼料摂取量が多く、成長速度が速いため、わずかな栄養利用性の差が飼料要求率や出荷体重に反映されやすくなります。トウモロコシ・大豆粕主体の標準的な配合でも、大豆粕由来のマンナンや可溶性非デンプン性多糖が腸管内容物の性状に影響する可能性があります。β-マンナナーゼ補給に関するブロイラーのメタ解析では、エネルギーおよび栄養素利用の改善可能性が評価されており、飼料設計におけるエネルギーマトリクスの考え方とも関連します[4]。
ブロイラーでの利点は、成長性能だけに限定されません。腸管内容物の粘性低下、未消化栄養素の後腸流入抑制、短鎖脂肪酸産生環境の変化、腸管バリアへの負荷軽減などが組み合わさることで、敷料の湿り、腸炎リスク、衛生状態にも間接的に関係し得ます。ただし、病原体制御や疾病予防を目的とする添加物ではないため、ワクチン、衛生管理、飼養密度、換気、敷料管理とは切り分けて評価する必要があります。ブロイラー腸内病原体の定着に関する栄養介入レビューでも、腸内環境への影響は多因子的であることが示されています[6]。
採卵鶏では、産卵率、卵重、卵殻品質、飼料効率、腸内環境が長期的な収益性に直結します。産卵後期や換羽後では、消化管機能や栄養要求のバランスが変わり、飼料中の難消化性多糖が生産性能へ影響しやすくなる場合があります。換羽後採卵鶏を対象としたβ-マンナナーゼ飼料酵素の研究では、複数の白色卵系統における産卵成績が検討されており、採卵鶏でもマンナン分解酵素が実務上の検討対象になっています[8]。
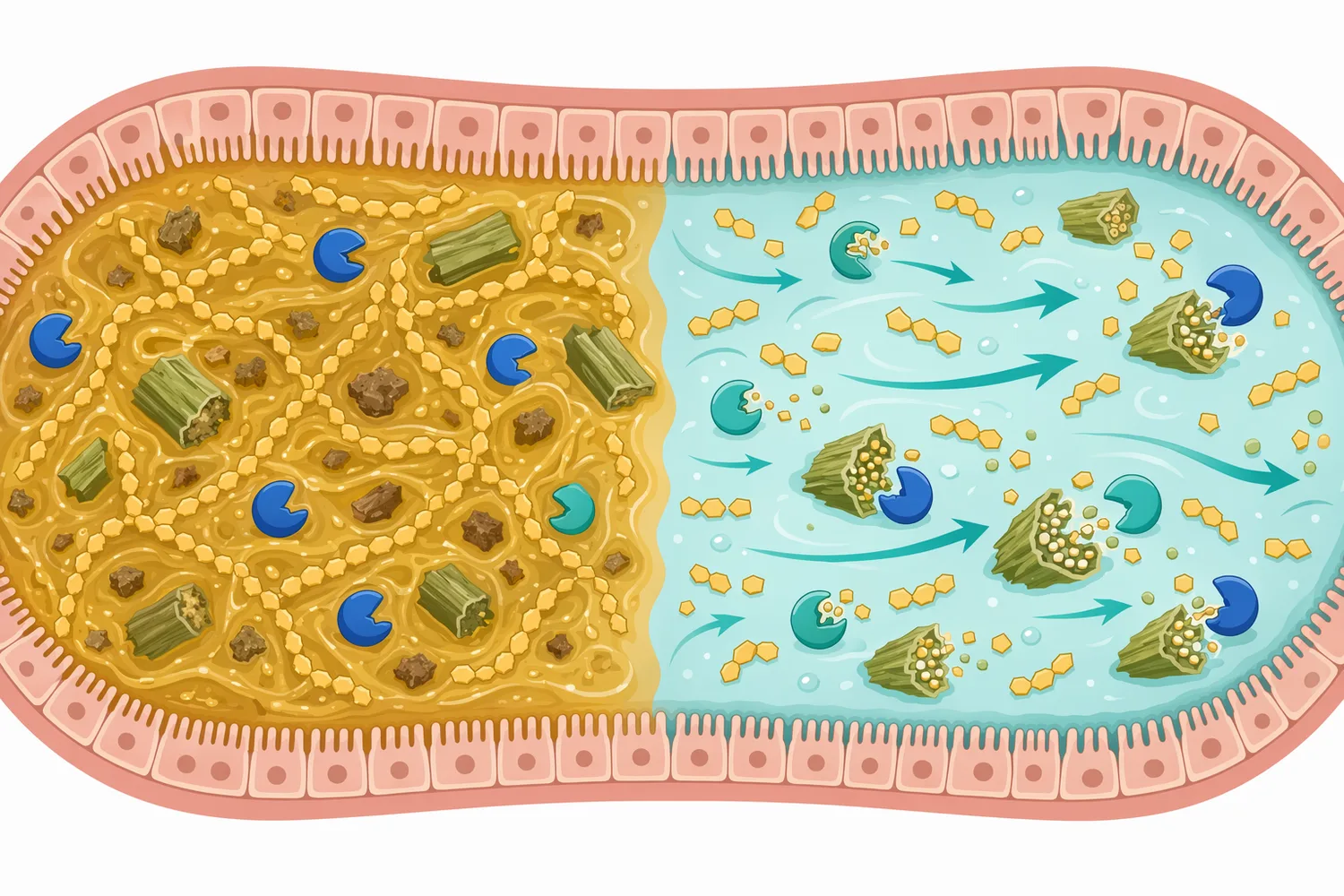
採卵鶏では、マンナナーゼを卵品質改善剤として単純に位置づけるよりも、植物性原料の利用性を高め、消化管負荷を抑え、産卵期の栄養供給を安定させる添加物として説明する方が正確です。卵殻品質や卵内容品質はカルシウム、リン、ビタミンD、微量ミネラル、熱ストレス、疾病、日齢などの影響が大きく、マンナナーゼ単独で一貫した改善を期待するのは適切ではありません。採卵鶏における天然・合成飼料添加物の研究でも、免疫や卵品質は複数要因の組み合わせとして評価されています[9]。
離乳子豚では、飼料の変化、母乳から固形飼料への移行、胃酸分泌の未成熟、腸内微生物叢の再編成、社会的ストレスが重なります。この時期に大豆粕など植物性タンパク質原料を多用すると、マンナン系多糖や抗栄養成分が腸管に負荷をかける可能性があります。離乳子豚を対象としたβ-マンナナーゼの研究では、異なる飼料タイプにおける腸管健康と成長への栄養的・機能的役割が検討されており、若齢豚での利用意義が明確に取り上げられています[1]。
離乳子豚で期待されるのは、下痢を直接止める薬理作用ではなく、消化しにくいマンナンを減らして腸管内の発酵基質と免疫刺激を調整することです。未消化タンパク質や炭水化物が後腸に過剰に流れると、浸透圧、発酵産物、pH、病原菌増殖のバランスが崩れやすくなります。β-マンナナーゼは、飼料原料由来のマンナン負荷を小さくすることで、離乳期の腸内環境を支える栄養管理ツールとして扱うのが妥当です[1]。
育成・肥育豚では、飼料費が生産コストの大部分を占めるため、原料の栄養価をどれだけ引き出せるかが重要です。トウモロコシ・大豆粕飼料、または副産物を含む飼料では、β-マンナンを含む非デンプン性多糖が消化率とエネルギー利用に影響します。全肥育期間にわたるβ-マンナナーゼ添加研究では、生産歩留まりと腸管健康が評価されており、長期給与での実務的検討が行われています[7]。
一方、豚では指標によって結果が一貫しないこともあります。成長成績、糞便性状、腸内細菌、炎症指標、枝肉形質は、基礎飼料、衛生状態、遺伝系統、日齢、飼育密度、疾病圧の影響を大きく受けます。育成豚でβ-マンナナーゼ単独またはマルチカルボヒドラーゼ複合体と組み合わせた研究は、栄養代謝と腸管健康の両面を扱っていますが、酵素添加の効果は飼料中の基質量と他酵素との相互作用を踏まえて解釈する必要があります[5]。
| 飼料原料・配合条件 | β-マンナン関連の特徴 | マンナナーゼで狙う主な作用 | 解釈上の注意 |
|---|---|---|---|
| 大豆粕主体の鶏・豚飼料 | 植物性タンパク質と非デンプン性多糖を同時に含む | 腸管粘性、未消化栄養素、免疫負荷の低減支援 | 標準的な原料でも効果は基質量と日齢に依存 |
| パーム核粕配合飼料 | マンナン系多糖が多く、難消化性が課題になりやすい | 細胞壁構造の分解、エネルギー利用性の補助 | 原料ロット差と加工条件の影響が大きい |
| コプラミール配合飼料 | マンナン含量が高く、副産物利用時の制約になりやすい | 副産物原料の栄養改善、消化性支援 | 高配合では他の栄養制約も同時に評価が必要 |
| 若齢動物向け高栄養飼料 | 消化管が未成熟で抗栄養因子の影響を受けやすい | 離乳期・初期成長期の腸管負荷軽減 | 衛生・温度・飼料摂取量の影響が大きい |
| マルチエンザイム配合飼料 | キシラン、グルカン、フィチン酸など複数基質が存在 | マンナン分解を他酵素作用と補完 | 基質の重複と相互作用を考慮する必要 |
コプラミールやパーム核粕のような副産物原料では、マンナン系多糖が栄養価を制限する主要因になりやすく、マンナナーゼの作用基質が明確です。コプラミールの栄養改善を扱った研究では、マンナナーゼと酵母の利用により副産物原料の価値向上が検討されており、マンナン分解が副産物活用の鍵になることが示されています[3]。
一方、大豆粕主体の標準飼料では、マンナン含量が副産物原料ほど高くない場合でも、若齢動物や高生産性動物では小さな消化性差が成績に影響することがあります。ブロイラーのメタ解析や離乳子豚の研究が示すように、β-マンナナーゼは「特殊原料専用」ではなく、植物性飼料全般のマンナン負荷を調整する酵素として使われます[4]。
マンナナーゼは、キシラナーゼ、β-グルカナーゼ、フィターゼ、プロテアーゼ、アミラーゼと同じ「外因性酵素」ですが、標的基質が異なります。キシラナーゼはアラビノキシラン、β-グルカナーゼはβ-グルカン、フィターゼはフィチン酸、プロテアーゼはタンパク質、アミラーゼはデンプンを主に対象とします。したがって、マンナナーゼの効果はβ-マンナンが存在する飼料で最も説明しやすく、他の酵素で完全に代替できるものではありません[2]。
| 酵素 | 主な標的基質 | 飼料上の主な目的 | マンナナーゼとの関係 |
|---|---|---|---|
| マンナナーゼ | β-マンナン、ガラクトマンナン、グルコマンナン | 粘性低減、マンナン抗栄養作用の緩和、腸内環境支援 | 大豆粕・パーム核粕・コプラミール利用時に重要 |
| キシラナーゼ | アラビノキシラン | 小麦・ライ麦・副産物由来の粘性対策 | 穀類由来NSP対策として補完的 |
| β-グルカナーゼ | β-グルカン | 大麦・オート麦由来の粘性対策 | 穀類配合で補完的 |
| フィターゼ | フィチン酸 | リン利用性改善、ミネラル結合の緩和 | 炭水化物分解ではなくリン栄養が中心 |
| プロテアーゼ | 飼料タンパク質 | アミノ酸利用性支援 | タンパク質消化を直接補助 |
| アミラーゼ | デンプン | エネルギー利用性支援 | デンプン消化を直接補助 |
マルチエンザイム製剤の研究では、複数の基質を同時に狙う設計が成長性能、栄養消化率、腸内微生物に影響し得ることが示されています。たとえば離乳子豚でトウモロコシ・小麦または小麦・大麦主体飼料に新規マルチエンザイムを添加した研究では、成長性能、栄養消化率、腸内マイクロバイオームが評価されています[10]。ただし、マルチエンザイムであっても、各酵素の効果は基質の有無に依存します。
マンナナーゼを他酵素と組み合わせる場合、効果は単純な足し算とは限りません。キシラナーゼやβ-グルカナーゼが粘性を下げ、マンナナーゼがマンナンを分解し、フィターゼがリン利用性を改善するように、作用点は異なります。育成豚でβ-マンナナーゼ単独またはマルチカルボヒドラーゼ複合体との併用を検討した研究では、栄養代謝と腸管健康が評価されており、酵素間の補完性を飼料基質に合わせて考える必要があります[5]。
最も根拠が明確なのは、β-マンナナーゼがβ-マンナンを加水分解し、植物性飼料中の抗栄養的なマンナン負荷を下げるという点です。これは酵素と基質の関係が明確で、飼料用外因性酵素の一般原理にも合致します。外因性酵素のレビューでは、酵素添加物の有効性は標的基質、消化管内での作用、飼料組成に依存すると整理されており、マンナナーゼは基質特異性の観点から説明しやすい酵素です[2]。

ブロイラーにおけるエネルギー・栄養素利用も、比較的検討が進んだ領域です。β-マンナナーゼ補給に関するメタ解析では、飼料中のエネルギーおよび栄養素利用が評価されており、個別試験のばらつきを統合して解釈する枠組みが示されています。これは、マンナナーゼの価値を「成長促進」という曖昧な表現ではなく、「マンナン分解を通じた栄養利用性支援」として説明する根拠になります[4]。
離乳子豚の腸管健康も重要な対象です。離乳後は腸管が不安定で、飼料由来の抗栄養因子に敏感なため、β-マンナン分解による粘性低減、未消化基質の減少、免疫負荷の調整が実務的に意味を持ちます。離乳子豚を対象にβ-マンナナーゼの栄養的・機能的役割を検討した研究は、飼料タイプによって反応が変わる点も含めて、現場での解釈に有用です[1]。
腸内微生物叢への影響は有望ですが、条件依存性が大きい領域です。マンナン分解物が発酵基質になり、短鎖脂肪酸産生や乳酸菌群に影響する可能性はありますが、同じ酵素でも飼料組成、飼育環境、抗菌剤使用、疾病圧によって菌叢変化は異なります。ブロイラー腸内の病原体定着に関する栄養介入レビューは、飼料添加物が腸内微生物に影響し得る一方、結果の解釈には多因子性を考慮すべきことを示しています[6]。
豚の育成・肥育期における腸管健康や生産成績も、中程度の根拠として扱うのが妥当です。全肥育期間でβ-マンナナーゼ添加を検討した研究では、生産歩留まりと腸管健康が評価されていますが、豚では飼料、遺伝、飼養管理、疾病状態の影響が大きく、すべての環境で同じ反応が出るとは限りません[7]。
枝肉品質、肉質、卵品質、疾病予防、病原菌制御などは、マンナナーゼ単独の直接効果として断定しにくい領域です。これらの指標は、栄養、遺伝、飼養環境、ストレス、感染、処理条件などの影響を受けます。飼料添加物全般の研究でも、免疫や生産物品質の反応は添加物の種類、投与条件、動物状態により大きく変わることが示されています[9]。

また、マンナナーゼはマイコトキシン吸着剤、抗菌剤、酸化防止剤、ワクチン、治療薬の代替ではありません。飼料中のアフラトキシン対策を扱った系統的レビューでは、毒素低減には目的に応じた添加物の選択と評価が必要であることが示されていますが、マンナナーゼの標的はあくまでマンナン系多糖です[11]。したがって、毒素、感染症、熱ストレスなどをマンナナーゼで直接解決できるような表現は避けるべきです。
マンナナーゼは、配合飼料の栄養設計において「原料の見かけの制限要因を下げる」ための酵素です。β-マンナンを分解することで、飼料原料に閉じ込められた栄養素を利用しやすくし、消化管内容物の物性を改善し、腸管免疫や微生物発酵への負荷を軽減する方向に働きます。外因性酵素のレビューでは、飼料酵素は動物の内因性消化だけでは十分に利用できない成分を対象にする添加物として整理されています[2]。
実際の効果は、飼料中にどれだけβ-マンナン基質があるかに左右されます。副産物原料を多く使う飼料、大豆粕比率が高い飼料、若齢動物向け飼料では、マンナン分解の余地が大きくなります。一方、マンナン含量が低い飼料、すでに消化性が高い原料構成、疾病や環境ストレスが主因の成績低下では、マンナナーゼの寄与は限定的になる可能性があります。コプラミールの栄養改善研究は、基質が明確な場合にマンナナーゼの利用意義が高まることを示す例です[3]。
マンナナーゼを「抗菌剤代替」と表現する場合も注意が必要です。抗菌性成長促進剤の使用制限下で、腸内環境を支える栄養技術の一つとしては有用ですが、抗菌作用そのものを持つわけではありません。ハーブ由来添加物や植物性添加物の研究では、抗菌剤代替として多様な添加物が検討されていますが、それぞれ作用機序は異なります[12]。マンナナーゼの場合、中心はマンナン分解とそれに伴う消化管環境の変化です。
Enzymes.bioは、動物飼料添加物向けマンナナーゼをオンラインで供給するサプライヤーです。製品は1kg単位で直接購入でき、注文処理に合わせてCoAおよびSDSが提供されます。Enzymes.bioは製造業者または研究機関としてではなく、B2B用途の酵素をオンラインで入手しやすくする供給者として位置づけられます。
本品は、飼料製造、畜産栄養、配合設計、研究開発の実務担当者が、β-マンナンを含む植物性原料の利用性を支援する目的で検討する酵素です。製品説明では、過度な成績改善の約束ではなく、β-マンナン分解、粘性低減、栄養素アクセスの改善、腸管環境支援という作用機序に沿って理解することが重要です。飼料用酵素の有効性は標的基質と配合条件に依存するため、マンナナーゼも「マンナン系多糖を含む飼料で意味を持つ添加物」として説明するのが科学的に正確です[2]。
マンナナーゼ酵素は、動物飼料中のβ-マンナン、ガラクトマンナン、グルコマンナンを加水分解し、植物性原料の消化利用性と腸内環境を支援する飼料添加物です。特にブロイラー、採卵鶏、離乳子豚、育成・肥育豚では、β-マンナンが粘性、未消化栄養素、免疫負荷、腸内微生物叢に影響し得るため、マンナナーゼの作用機序は実務的に説明しやすいものです[1]。
科学的に最も確かな説明は、β-マンナナーゼがβ-1,4-マンナン結合を切断し、マンナン系抗栄養因子を低分子化するという点です。ブロイラーではエネルギー・栄養素利用、離乳子豚では腸管健康、育成・肥育豚では栄養代謝と腸内状態への影響が研究されており、飼料原料の組成と動物の生理段階に応じて価値が変わります[4]。
一方で、マンナナーゼは万能な生産性改善剤ではありません。枝肉品質、卵品質、疾病予防、マイコトキシン対策などを直接保証する添加物ではなく、β-マンナンを含む飼料原料の利用性を高めるための基質特異的な酵素です。Enzymes.bioのマンナナーゼは、1kg単位でオンライン購入できる動物飼料添加物向け酵素として、マンナン系多糖を含む植物性飼料設計における消化性・腸内環境支援に適した選択肢です。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Mannanase Enzyme For Animal Feed Additives ≥10000U/Gを購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。