Mannanase feed addition helps break down β-mannan hemicellulose in plant-based feed materials into shorter mannose-rich oligosaccharides. In practical terms, it targets a specific non-starch polysaccharide fraction that can limit nutrient access, increase digesta viscosity, and contribute to anti-nutritional effects in some poultry, swine, and other feed systems.
For buyers using Enzymes.bio, Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processing is available for direct online purchase by the 1 kg unit. Payment is completed online, the order is processed and shipped, and a Certificate of Analysis and Safety Data Sheet are provided with the order.
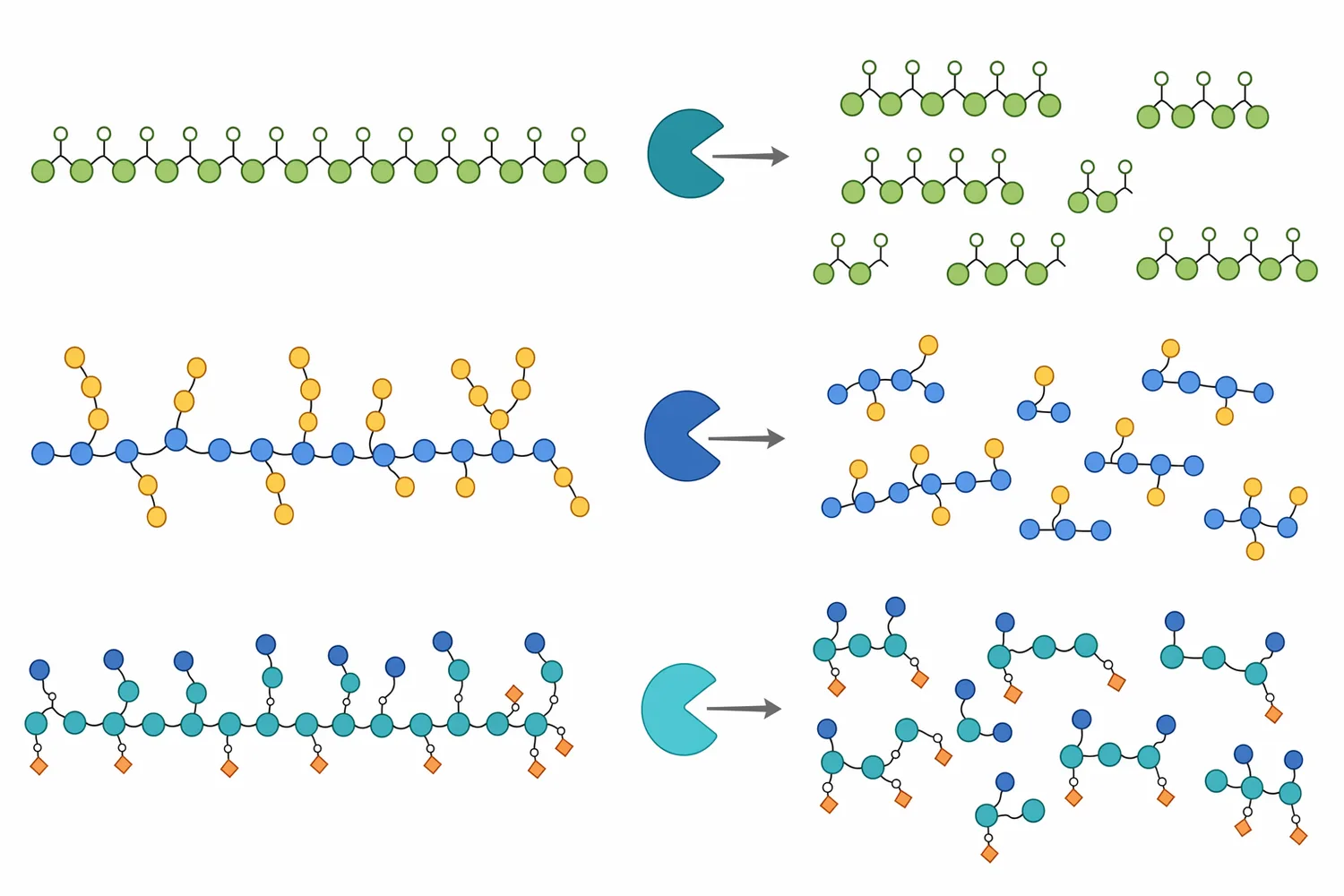
Mannanase is a carbohydrase that hydrolyzes mannan-based polysaccharides, especially β-1,4-linked mannan backbones found in mannans, galactomannans, glucomannans, and galactoglucomannans. These polymers are part of the hemicellulose fraction of plant cell walls, meaning they sit alongside cellulose, pectin, lignin-associated structures, arabinoxylans, and other non-starch polysaccharides in many feed ingredients and agricultural co-products. Hemicellulose is not a single molecule; it is a family of branched and substituted polysaccharides, and mannanase is useful because it addresses the mannose-containing part of that family rather than fiber in general [1].
In animal feed, β-mannans matter because they are common in vegetable raw materials and can be poorly utilized by monogastric animals. Soybean meal, palm kernel cake, palm kernel expeller, copra-derived materials, and several other plant co-products may contribute mannan-rich fractions to the final ration. The nutritional effect depends on inclusion level, solubility, molecular structure, and how accessible the mannan is after grinding, pelleting, extrusion, or other processing; however, the common technical issue is the same: long-chain β-mannans can remain intact through much of normal digestion unless an appropriate exogenous enzyme is present [2].
The “mannose oligosaccharide processing” part of the product description refers to what happens when the long polymer is cut into shorter fragments. Endo-β-mannanase does not simply dissolve plant material in a non-specific way. It attacks internal β-1,4 linkages in the mannan backbone, reducing polymer size and generating shorter mannose-containing oligosaccharides. Those smaller fragments behave differently from the original high-molecular-weight hemicellulose: they are more soluble, less structurally entangling, and less able to maintain the same cell-wall matrix effects that can restrict access to protein, starch, and lipids.
A useful way to understand mannanase is to picture the mannan polymer as a long, branched carbohydrate cable embedded in a plant cell-wall network. Before enzyme action, that cable can bind water, contribute to viscosity, physically obstruct digestive enzymes, and keep nutrients trapped inside fiber-rich particles. After mannanase action, the cable is cut into shorter sections. The physical network becomes less continuous, and the smaller fragments no longer create the same barrier to nutrient release.
At the molecular level, the enzyme binds along the mannan chain in a catalytic groove and positions the β-1,4 glycosidic bond for hydrolysis. Studies on mannanase action against galactoglucomannan show that substitution patterns such as acetylation influence how readily the enzyme can access and cleave the backbone; in other words, the enzyme is acting on a real plant polymer with side groups and branching, not an idealized straight chain [1]. This matters commercially because two ingredients can have similar crude fiber values but respond differently if one contains more accessible β-mannan and the other contains mannan protected by substitution, lignification, or processing history.
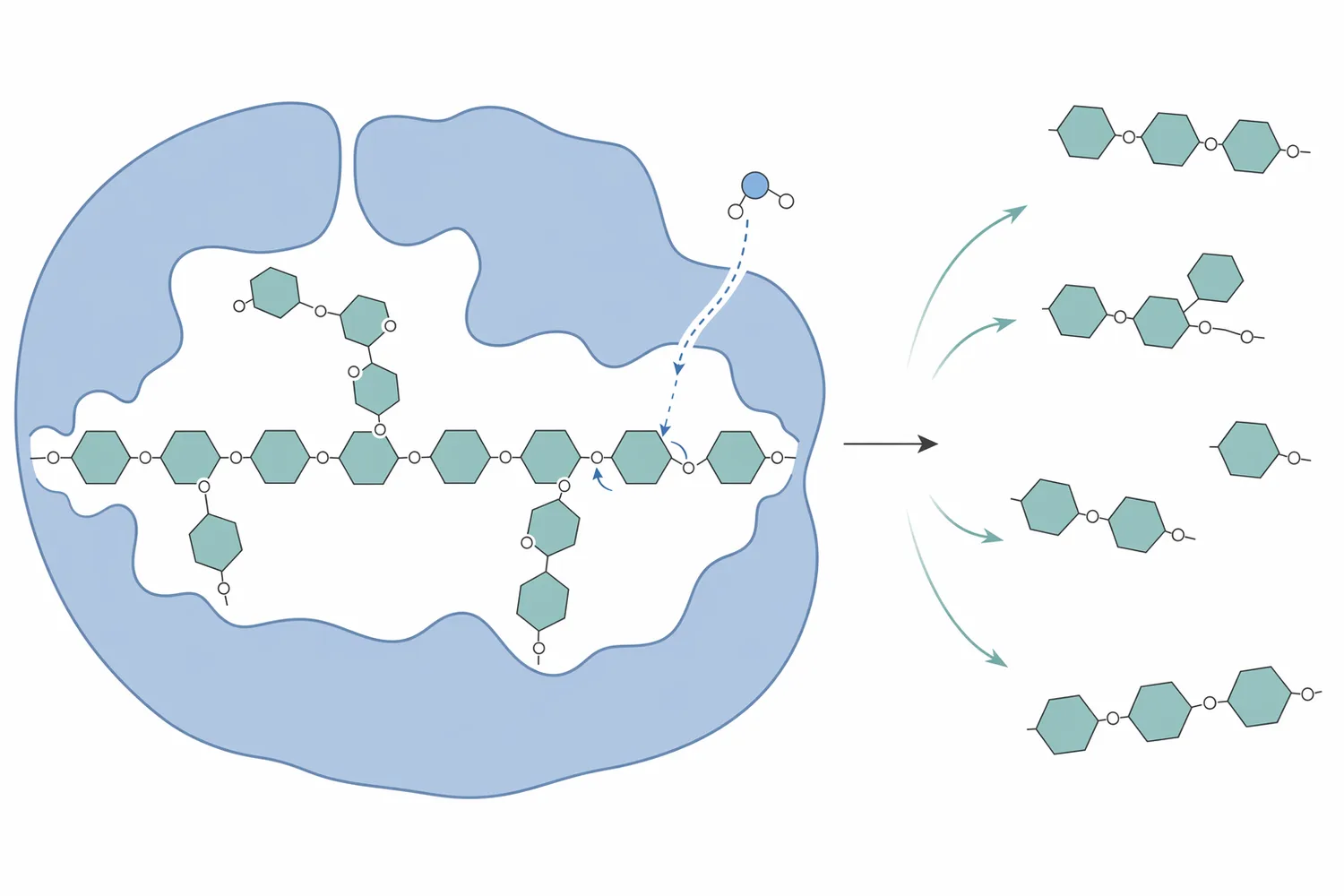
The result is not only a change in carbohydrate size. When the hemicellulose network is partially depolymerized, associated nutrients can become more available to the digestive process. Protein bodies, oil residues, and starch granules that were physically enclosed in plant tissue may be more exposed. In palm kernel cake and related co-products, where mannan-rich fiber is a major limitation, studies have examined pretreatment and mannanase hydrolysis specifically because the ingredient’s carbohydrate structure can restrict its feed value [3].
| Feed or ingredient feature | Before mannanase action | After β-mannanase hydrolysis | Practical meaning |
|---|---|---|---|
| β-mannan chain length | Long mannose-rich backbone remains largely intact | Backbone is cleaved into shorter mannose-rich oligosaccharides | Lower structural persistence of the mannan fraction |
| Cell-wall matrix | Hemicellulose helps hold plant particles together | Matrix is partially loosened where mannan is accessible | More opportunity for digestive enzymes to contact nutrients |
| Soluble fiber behavior | Soluble β-mannans may contribute to viscosity and anti-nutritional effects | Smaller fragments have different physical behavior | Reduced impact of high-molecular-weight mannan structures |
| Nutrient access | Protein, fat, and carbohydrates may remain trapped in fiber-rich particles | Enzyme action can expose entrapped nutrients | Better use of mannan-containing plant ingredients where conditions are suitable |
| Oligosaccharide profile | Larger polysaccharides dominate | Mannose-containing oligosaccharides increase | More processed hemicellulose fraction |
The term “anti-nutritional” does not mean β-mannans are toxic by default. It means that, in certain diet contexts, they can reduce the efficiency with which animals use the nutrients already present in the ration. This can happen through physical effects, such as increased viscosity and reduced diffusion of digestive enzymes, and through matrix effects, where nutrients are trapped within plant cell-wall structures. It may also involve physiological responses to soluble β-mannan fractions in the gut.
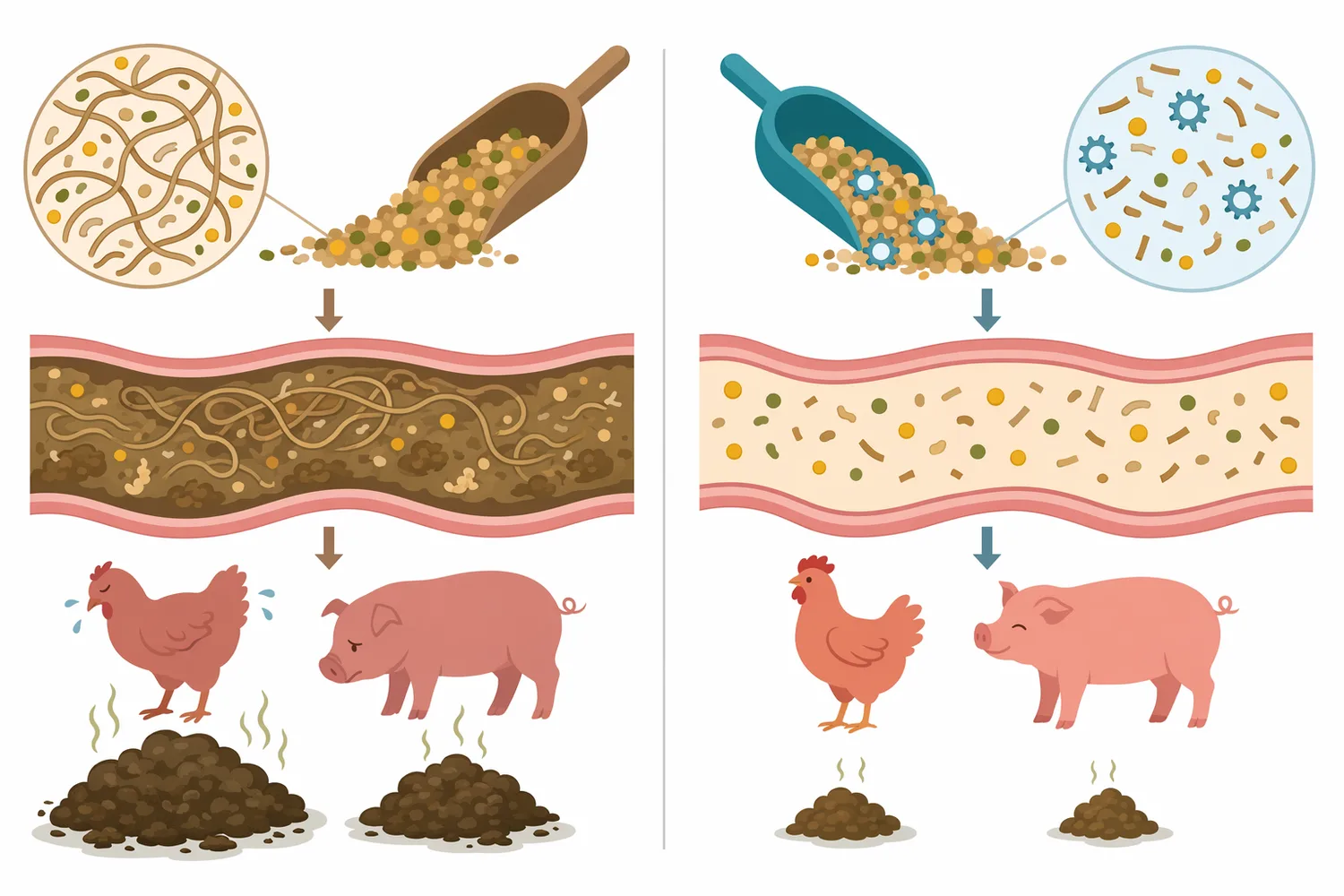
A major body of feed-enzyme research has examined β-mannanase in broiler chickens, laying hens, turkeys, sows, and nursery-to-finish pigs, reflecting how widely the substrate occurs across practical livestock diets [2]. The relevance is especially strong for monogastric animals, because pigs and poultry do not produce enough endogenous enzymes to efficiently depolymerize many non-starch polysaccharides. Their digestive systems can use starch, protein, and fat efficiently when those nutrients are accessible, but intact hemicellulose can limit that access.
Nutrition researchers also discuss a “feed-induced immune response” associated with soluble β-mannans. The proposed mechanism is that some soluble β-mannan structures may be recognized in ways that activate innate immune pathways, diverting energy and nutrients toward immune activity rather than growth or production. A 2022 appraisal specifically reviewed whether β-mannanase supplementation can modulate feed-induced immune response and gastrointestinal ecology in poultry and pigs, indicating that this is an active and practically relevant research area rather than a marketing invention [4].
This mechanism should be handled carefully. It supports a plausible biological explanation for some performance responses, but it does not mean every mannan-containing diet will produce the same immune effect or that enzyme addition should be described as a health treatment. The most robust customer-facing statement is narrower and stronger: mannanase reduces the size and structural effect of β-mannan hemicellulose, which can support nutrient utilization and gut-function conditions when β-mannans are a meaningful limiting factor in the diet.
Poultry diets are one of the best-studied areas for exogenous carbohydrases because broilers and turkeys are highly sensitive to feed conversion, gut viscosity, and the digestibility of plant-based ingredients. A systematic review and meta-analysis evaluated single β-mannanase supplementation across broiler chickens, laying hens, turkeys, sows, and pigs, focusing on performance and energy utilization outcomes [2]. The inclusion of multiple poultry categories is important because it shows that β-mannanase is not limited to one production stage or one feed ingredient, even though responses remain diet-dependent.
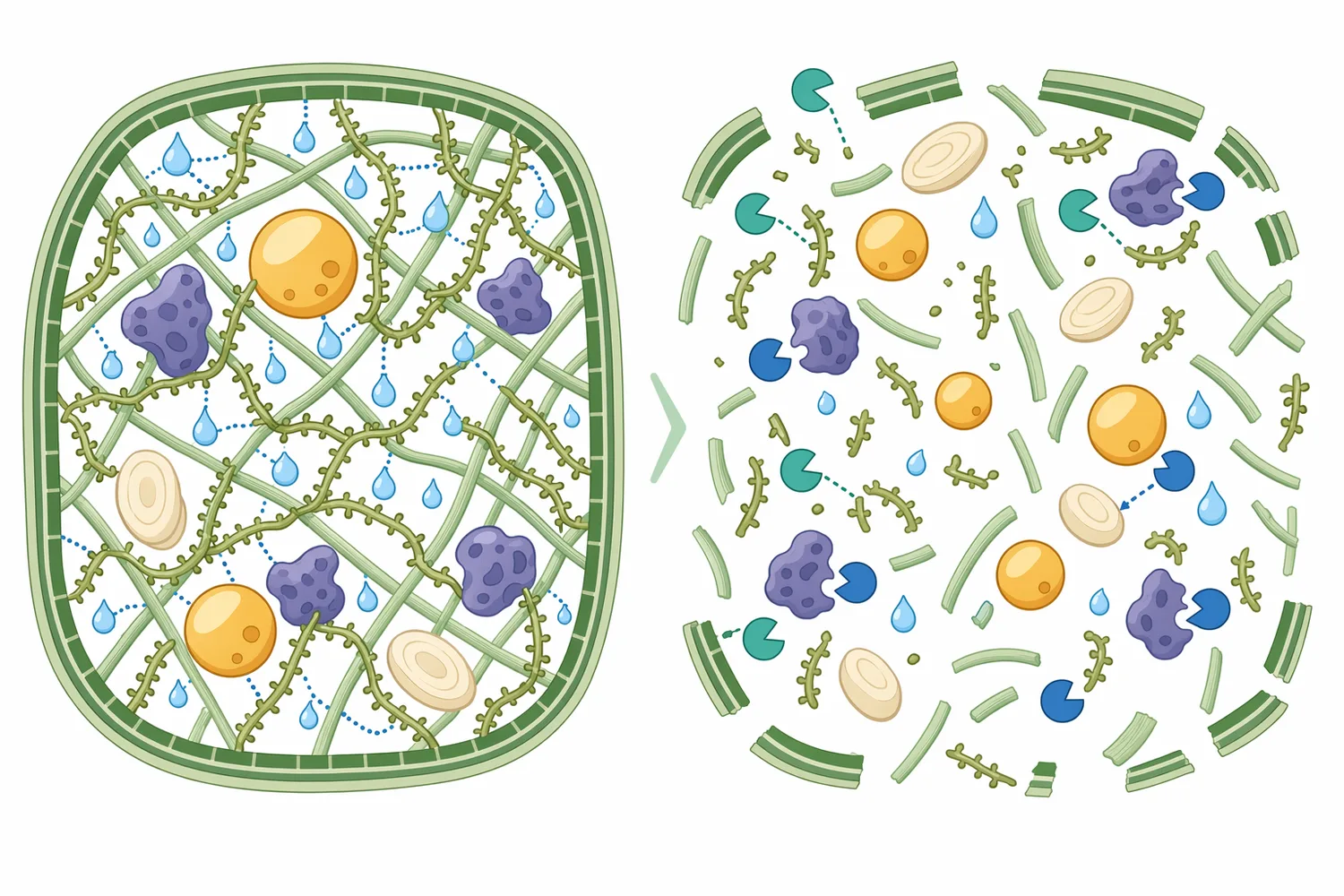
More recent broiler work has looked at β-mannanase alone and in combination with xylanase and β-glucanase in wheat-based diets. That design is technically meaningful because wheat-based rations contain multiple non-starch polysaccharide fractions, not only mannans; xylanase addresses arabinoxylans, β-glucanase addresses β-glucans, and β-mannanase addresses mannans. The 2025 broiler study reported enhanced growth performance, non-starch polysaccharide degradation, and gastrointestinal environment when β-mannanase was used alone or in enzyme combinations in wheat-based diets [5].
This type of evidence helps explain the role of mannanase in multi-enzyme feed strategies. Mannanase is not a substitute for every other enzyme. It does not hydrolyze phytate like phytase, does not hydrolyze protein like protease, and does not specifically target arabinoxylan like xylanase. Its value is precision: it addresses the β-mannan portion of the hemicellulose burden, which may be one of several fiber barriers in a complex plant-based ration.
Low-energy diet studies also help place mannanase in commercial context. Broiler research has evaluated β-mannanase separately and in combination with a cocktail non-starch polysaccharide enzyme system in low-energy diets, measuring performance and processing-related parameters [6]. The practical logic is that if an enzyme improves the release or utilization of nutrients already present in the diet, it may help animals maintain performance under formulations designed to reduce feed cost or energy density. That does not make energy reduction automatic; it shows why enzyme-responsive fiber fractions are important when diets are reformulated.
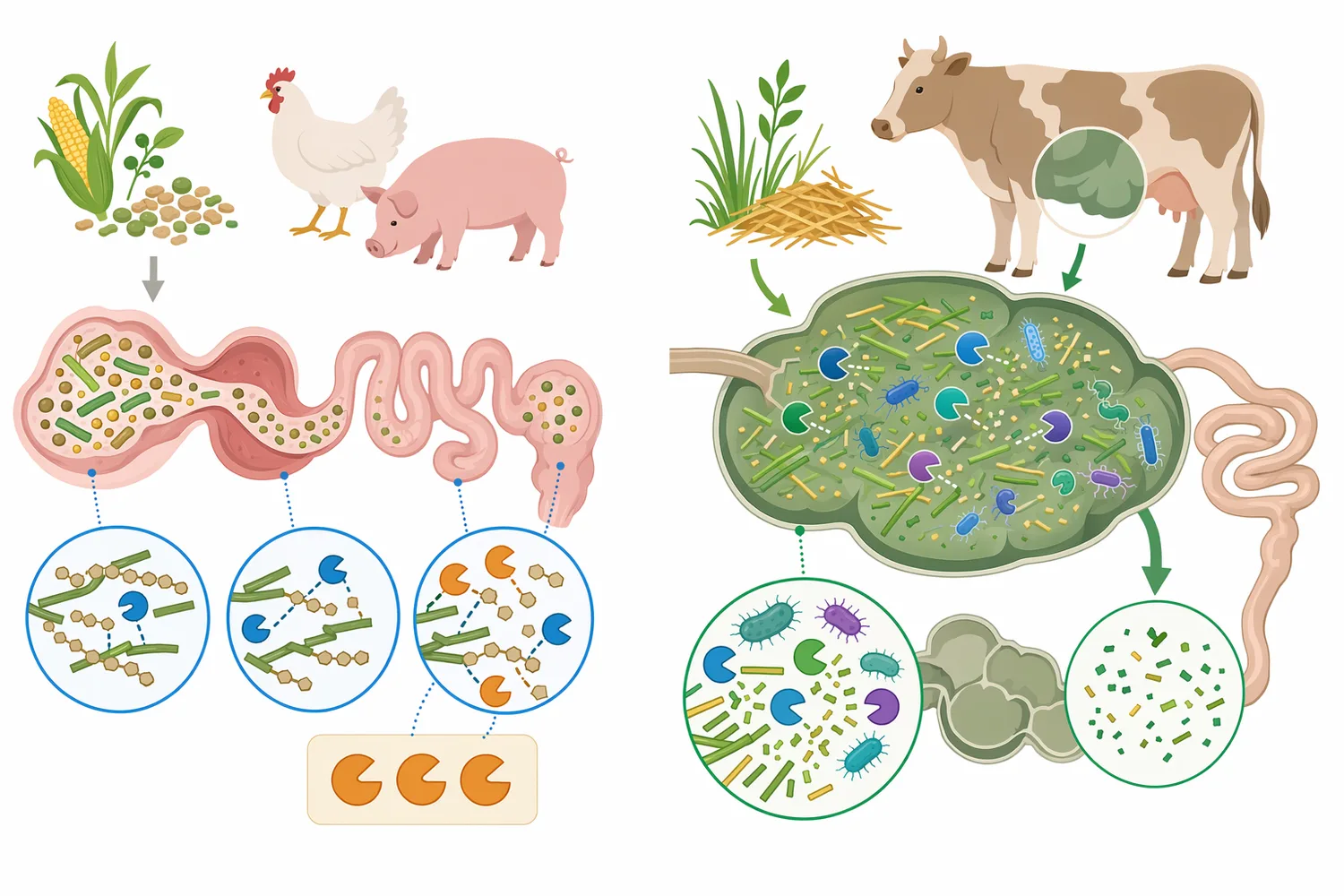
Swine diets often include soybean meal and may include palm kernel expeller, distillers grains, or other vegetable co-products depending on region and cost. Newly weaned pigs are especially sensitive to diet changes because their digestive capacity, gut barrier function, and microbiota are still adapting. Research on newly weaned pigs has investigated the nutritional and functional roles of mannanase in both typical and low-cost formulated diets, with emphasis on intestinal health and growth [7].
The mechanism in pigs is similar to poultry but expressed through a different digestive system. Intact β-mannan can increase the fiber burden in the small intestine and reduce access to nutrients before fermentation occurs later in the gut. By cutting β-mannan into smaller oligosaccharides, mannanase changes the substrate reaching different gut segments. This can influence the balance between nutrient digestion in the small intestine and microbial fermentation in the hindgut, which is why swine studies often consider both performance and intestinal environment.
The 2021 systematic review and meta-analysis is useful here because it included sows and nursery-finish pigs alongside poultry categories [2]. That breadth matters for buyers comparing mannanase with more familiar enzymes: the feed industry has long used phytase and xylanase, but β-mannanase has its own evidence base across production animals where mannan-rich ingredients are present. The expected value is highest when the formulation contains enough accessible β-mannan for the enzyme to act on.
For pig feeds using economical co-products, the benefit is often framed as improved utilization rather than simple nutrient addition. Mannanase does not add protein or energy by itself. Instead, it can make part of the existing feed matrix less resistant to digestion, so the animal can use more of the nutrients already purchased in the ration. That distinction is important: the enzyme acts as a processing and digestive aid for a specific fiber substrate, not as a replacement for balanced formulation.
Palm kernel cake and palm kernel expeller are among the clearest practical examples of why mannanase is used. These co-products can be attractive feed ingredients, but their high fiber and mannan content can limit inclusion if the carbohydrate matrix is not addressed. Research on selective hemicellulose degradation and mannan-rich biomass highlights that breaking hemicellulose into oligosaccharides changes the functionality of the material, making the carbohydrate fraction more processable [3].
In palm-derived co-products, the core problem is accessibility. The feed may contain valuable residual protein, oil, and energy, but those nutrients are partly embedded in a tough cell-wall matrix. Mannan-rich hemicellulose contributes to that matrix. When mannanase cleaves the β-mannan backbone, it can increase soluble carbohydrate fragments and reduce the integrity of the fiber barrier. Physical pretreatments such as heat, moisture, or particle-size reduction may also change accessibility, although the enzyme’s actual target remains the β-mannan linkage.
From a feed-processing viewpoint, this is why mannanase is often discussed as an ingredient-upgrading tool. It can help convert a co-product from “fiber-limited” toward “more digestible and more predictable,” provided the substrate contains accessible β-mannan. The enzyme’s effect will not be identical in every palm kernel source because oil extraction method, shell contamination, drying history, and particle structure can all affect how much of the mannan backbone is available for hydrolysis.
The practical output of mannanase treatment is a shift in the carbohydrate profile: less intact mannan-rich hemicellulose and more smaller mannose-containing fragments. In feed terms, that may support better nutrient release and reduced anti-nutritional effect. In ingredient-processing terms, it may make the co-product easier to integrate into formulations that rely on vegetable raw materials without overloading the animal with poorly digestible fiber.
Mannose oligosaccharides are short carbohydrate chains containing mannose units. They can arise when mannanase cuts longer mannan, glucomannan, galactomannan, or galactoglucomannan chains. The exact mixture depends on the substrate: a relatively linear mannan can yield different fragments from a branched galactomannan, and an acetylated galactoglucomannan can resist or redirect cleavage compared with a less substituted polymer [1].
This is why “mannose oligosaccharide processing” should be understood as a controlled direction of change rather than one single molecule. The enzyme reduces polymer size and shifts the distribution from high-molecular-weight hemicellulose toward shorter oligosaccharides. Some fragments may contain only mannose; others may include glucose, galactose, or acetyl substitutions depending on the original feedstock. In real feed materials, especially co-products, the output is a family of related oligosaccharides.
Recent enzyme research also shows that substrate branching can influence oligosaccharide release patterns. A rumen-derived bifunctional glucanase/mannanase was reported to release oligosaccharides with a high degree of polymerization preferentially from branched substrates, illustrating that enzyme architecture and substrate structure together determine what fragments are produced [8]. For feed applications, the main takeaway is practical: the same enzyme category can behave differently on soybean galactomannan, palm kernel mannan, or softwood galactoglucomannan because the polymer architecture differs.
The oligosaccharide result is relevant to animal nutrition because shorter fragments no longer behave like the original intact fiber. They may be more fermentable, less viscous, and less able to form the same barrier around nutrients. However, it is more accurate to describe the benefit as hemicellulose modification and support for feed utilization than to overstate it as a guaranteed prebiotic or health effect in all animals.
Feed enzyme systems are often discussed as a group, but each enzyme has a distinct substrate. Phytase targets phytate-bound phosphorus. Xylanase targets arabinoxylan. β-glucanase targets β-glucans. Protease targets proteins. Mannanase targets β-mannan hemicellulose. Understanding that distinction helps avoid unrealistic expectations: if the feed problem is mainly phytate, mannanase is not the primary enzyme; if the feed problem is mannan-rich fiber, mannanase becomes more relevant.
The 2025 broiler study combining β-mannanase with xylanase and β-glucanase illustrates this substrate-specific approach. In wheat-based diets, multiple non-starch polysaccharides can contribute to viscosity and nutrient encapsulation, so combining enzymes can broaden the range of hydrolyzed fiber structures [5]. In contrast, a diet dominated by palm kernel co-products may place more emphasis on mannanase because β-mannan is a major structural carbohydrate in that ingredient class.
| Enzyme type | Main substrate in plant-based feeds | Primary structural change | Feed relevance |
|---|---|---|---|
| Mannanase | β-mannans, galactomannans, glucomannans, galactoglucomannans | Cleaves mannose-rich hemicellulose backbones into shorter oligosaccharides | Supports processing of mannan-rich ingredients and co-products |
| Xylanase | Arabinoxylans and related xylans | Reduces arabinoxylan chain length and viscosity effects | Common in wheat, rye, and other cereal-based diets |
| β-glucanase | β-glucans | Breaks down β-glucan polymers | Relevant in barley, oats, and some cereal fractions |
| Phytase | Phytate | Releases phytate-bound phosphorus and reduces phytate anti-nutritional effects | Used widely to improve phosphorus availability |
| Protease | Feed proteins | Hydrolyzes protein chains into smaller peptides | Supports protein digestibility where protein accessibility is limiting |
This comparison also explains why mannanase is described as a feed addition rather than a complete feed solution. It performs one well-defined biochemical task. The commercial value comes from matching that task with ingredients where β-mannan is part of the nutritional bottleneck.
Although much of the feed-enzyme discussion focuses on monogastric animals, mannanase is also being explored in ruminant nutrition. A 2025 review examined β-mannanase supplementation in dairy cattle nutrition, performance, and environmental sustainability, showing that the enzyme is being considered beyond poultry and swine systems [9]. In ruminants, the rumen microbiome already has substantial fiber-degrading capacity, so the role of exogenous enzymes is different from monogastric digestion and must be interpreted in that context.
The ruminant connection is scientifically interesting because natural rumen microbes produce enzymes that degrade plant polysaccharides. Research on a rumen-derived bifunctional glucanase/mannanase highlights how microbial enzymes can attack branched substrates and release oligosaccharides with distinct size profiles [8]. This supports the broader biological principle that mannanase-type enzymes are part of natural fiber conversion systems, even though commercial feed addition must be evaluated according to the target species and diet.
Aquafeed is another area where plant ingredients and co-products are increasingly used, although the direct mannanase evidence base is more limited in the supplied literature. Reviews of natural feed supplements and processing side streams for finfish and crustaceans show strong interest in upgrading non-traditional ingredients, improving digestibility, and using co-products more effectively [10]. Where mannan-rich plant materials are used in aquafeed, the same substrate logic applies, but species-specific digestive physiology remains important.
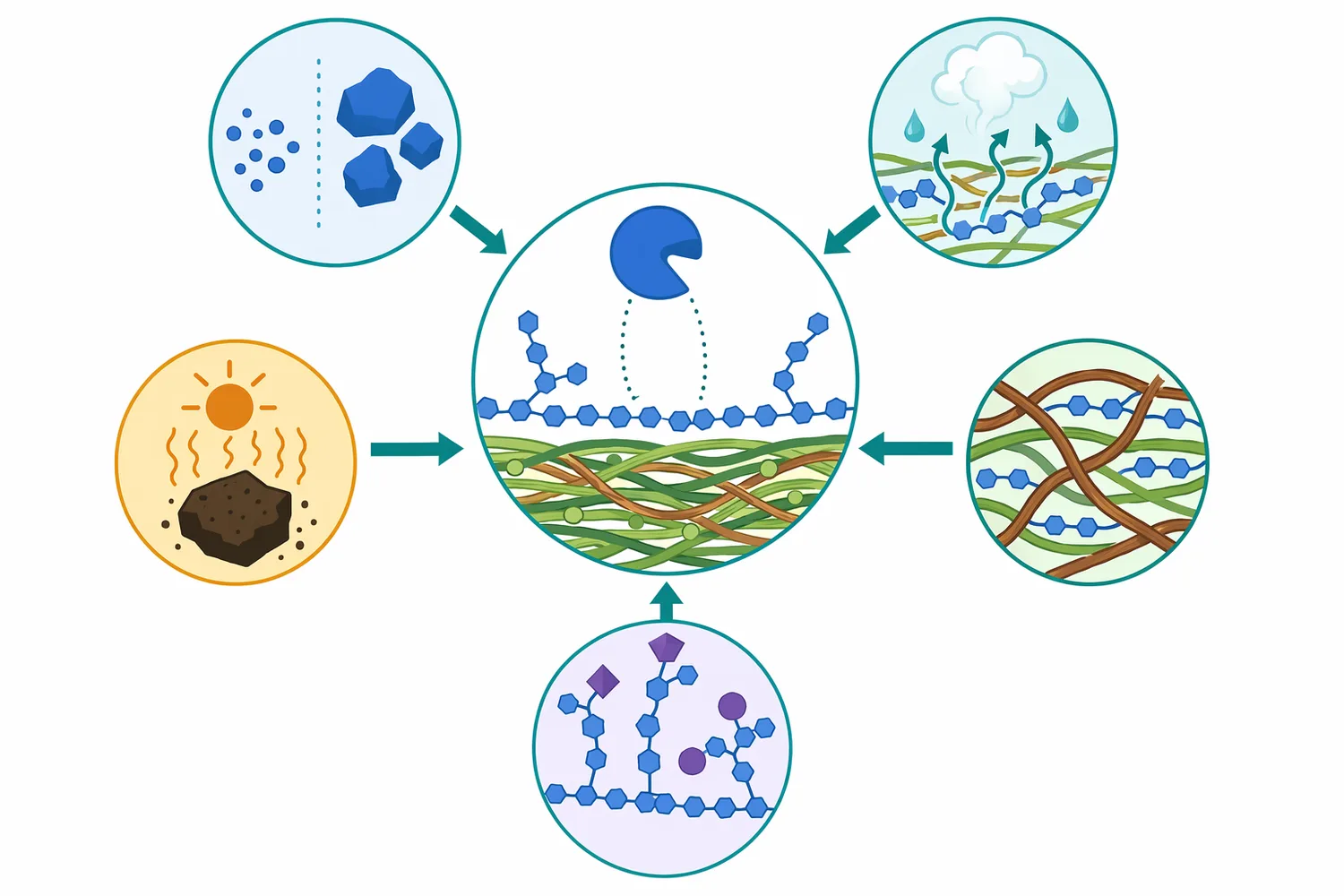
Enzyme performance depends on whether the enzyme can physically reach its substrate. In a finely ground soybean-derived material, soluble galactomannan may be more accessible than mannan embedded in a lignified, coarse, or heat-damaged particle. In palm kernel cake, steam or moisture processing may alter the cell-wall architecture, making some hemicellulose more available for hydrolysis. The key concept is not simply “more processing is better”; it is that physical structure controls how much β-mannan surface is exposed to the enzyme.
Studies on hemicellulose degradation in plant biomass repeatedly show that cellulose, hemicellulose, and lignin interact within the cell wall. Thermal and physical treatments can change the degradation behavior of hemicellulose and cellulose because these polymers are not isolated in the native material [11]. For feed applications, this means that the same mannanase can show different practical effects depending on particle size, pelleting history, moisture, heat exposure, and whether the feed material has already been processed into a more open structure.
Acetylation and branching are also important. Work on spruce galactoglucomannan demonstrated that acetylation affects enzymatic mannan degradation, meaning chemical substitutions on the hemicellulose backbone can influence hydrolysis [1]. In practical terms, mannanase acts most readily where the β-1,4 backbone is accessible and not overly shielded by side groups, crystallinity, lignin association, or physical encapsulation.
Temperature and pH tolerance are enzyme-specific properties, and the scientific literature includes mannanases adapted to different operating environments. For example, research has described a low-temperature-active and organic-solvent-stable mannanase isolated from an intestinal metagenome of Hermetia illucens, showing the diversity of mannanase enzymes that can exist in nature [12]. For a feed buyer, the main point is that mannanase is a functional enzyme class with variants adapted to different biological and processing environments, not one identical molecule in every product.
Feed enzymes are used because they act catalytically on feed substrates and are generally applied at low inclusion relative to the total diet. Safety evaluation remains important, especially for target animals, users handling the product, consumers of animal-derived foods, and the environment. The European Food Safety Authority has evaluated Hemicell®-L, an endo-1,4-β-mannanase feed additive, for chickens for fattening or reared for laying, turkeys for fattening or reared for breeding, and minor poultry species [13].
That type of evaluation does not automatically apply every conclusion to every mannanase product, but it does show that endo-β-mannanase as a feed-additive category has been examined in formal animal-feed safety and efficacy contexts. For commercial users, this supports confidence in the general enzyme approach while keeping claims tied to the product’s intended technical function: hydrolysis of β-mannan-containing hemicellulose.
The most responsible positioning is therefore neither exaggerated nor vague. Mannanase should not be presented as a medicine, growth promoter, or universal solution for poor feed quality. It should be presented as a targeted enzyme that modifies a known carbohydrate substrate. When that substrate is present and accessible, the enzyme can support more efficient use of plant-derived feed materials.
In poultry, mannanase is most relevant where soybean meal, palm-derived co-products, or other mannan-containing plant materials contribute to the non-starch polysaccharide load. The enzyme can reduce the structural and soluble effects of β-mannans, complementing other carbohydrases when cereal arabinoxylans or β-glucans are also present. Studies in broilers using β-mannanase alone or with xylanase and β-glucanase support the concept that targeted NSP degradation can improve the gastrointestinal environment and feed utilization in appropriate diets [5].
For pigs, mannanase is useful where cost-effective plant ingredients increase the mannan-rich fiber fraction. Newly weaned pigs may be particularly sensitive to fiber quality and nutrient accessibility, while grower-finisher pigs may benefit from improved utilization of co-products. Research examining mannanase in newly weaned pigs fed low-cost formulated diets reflects this practical interest in maintaining nutritional function while using economical raw materials [7].
Palm kernel cake and expeller are strong candidates for mannanase-supported processing because their fiber limitation is closely linked to mannan-rich hemicellulose. Enzyme action can shift the material toward smaller soluble carbohydrate fragments and may improve access to residual protein and energy. Research on hemicellulose degradation into oligosaccharides supports the technical principle that controlled breakdown changes material functionality rather than merely reducing fiber analytically [3].
Mannanase also has relevance in broader hemicellulose processing. Lignocellulosic biomass contains cellulose, hemicellulose, and lignin in complex networks, and mannans are one important hemicellulose class. Research into hemicellulose degradation, microbial biomass conversion, and mannanase substrate specificity helps explain why the enzyme is used in plant-material processing beyond direct animal nutrition [14].
A buyer should expect mannanase to perform a specific biochemical function: hydrolyzing accessible β-mannan hemicellulose into shorter mannose-rich oligosaccharides. That is the central claim supported by enzyme chemistry, plant cell-wall research, and animal-feed studies. The downstream benefits—improved nutrient access, reduced anti-nutritional effects, and better use of mannan-containing ingredients—are application-dependent because they rely on the diet actually containing accessible β-mannan in meaningful amounts.
The strongest evidence base supports three practical statements. First, β-mannanase is relevant across poultry and swine nutrition, with meta-analytic evaluation covering broiler chickens, laying hens, turkeys, sows, and nursery-to-finish pigs [2]. Second, mannanase can be part of multi-enzyme non-starch polysaccharide strategies, especially when diets contain more than one problematic fiber fraction [5]. Third, mannanase is especially logical for mannan-rich co-products, where the targeted substrate is a known contributor to limited digestibility and ingredient variability [3].
The cautious boundary is equally important. Mannanase does not make all fiber digestible, does not replace formulation expertise, and does not guarantee the same animal-performance response in every feed. If a diet contains little accessible β-mannan, the enzyme has less substrate to act on. If the mannan is physically shielded or chemically substituted in ways that limit access, hydrolysis may be reduced. If other nutrients are more limiting than fiber, the visible performance response may be modest even though the enzyme is functioning.
Enzymes.bio supplies Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processing as a direct online product sold by the 1 kg unit. Buyers place the order and complete payment online; the order is then processed and shipped. A Certificate of Analysis and Safety Data Sheet are provided with the order.
This purchasing model is designed for straightforward access to the enzyme without a lengthy quotation process. The product is best understood as a technical enzyme addition for feed and ingredient systems where β-mannan hemicellulose is a relevant substrate.
Mannanase feed addition is a targeted way to process β-mannan hemicellulose in plant-based feed materials. By cleaving β-1,4-linked mannan backbones, it converts large mannose-rich polymers into shorter oligosaccharides, reducing the structural persistence of the mannan fraction and helping expose nutrients trapped within plant cell-wall matrices.
The evidence base supports mannanase use in poultry, swine, and broader feed-enzyme systems, especially where soybean-derived ingredients, palm kernel cake, palm kernel expeller, or other mannan-containing co-products contribute to the non-starch polysaccharide load. Research also shows why responses vary: substrate branching, acetylation, processing history, diet design, and animal physiology all influence the practical outcome.
For buyers, the central value proposition is clear: Enzymes.bio Mannanase Feed Addition supports hemicellulose breakdown and mannose oligosaccharide processing in feed and ingredient applications where β-mannans are a meaningful limiting factor. It is a precise enzyme tool for improving the usability of mannan-rich plant materials, available for direct online purchase in 1 kg units with order documentation included.
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processing →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.