Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processingは、飼料中のβ-マンナン系ヘミセルロースを低分子化し、消化管内容物の粘性負荷を下げ、マンナン由来オリゴ糖の生成を支援するための飼料用途向けマンナナーゼです。対象は主に大豆粕、パーム核粕、コプラミール、その他の植物性副産物に含まれるマンナン、ガラクトマンナン、グルコマンナンであり、タンパク質や脂質を直接分解する汎用酵素ではありません。Enzymes.bioは本品を製造業者・研究所としてではなく、B2B向けオンライン酵素供給業者として1 kg単位で直接販売し、CoAおよびSDSは注文時に併せて提供されます。
飼料用マンナナーゼは、植物性原料の細胞壁・貯蔵多糖に由来するβ-マンナンを加水分解する外因性酵素です。β-マンナンはヘミセルロースの一群で、主鎖にβ-1,4結合したマンノース残基を持ち、原料によってはガラクトース側鎖、グルコース共重合、アセチル化などの構造差を伴います。マンナンは植物細胞壁の構造多糖としてだけでなく、種子内の貯蔵多糖としても存在し、植物種・組織・加工履歴によって可溶性、分岐度、酵素アクセス性が変わることが整理されています[1]。
本品の主要用途は、飼料中ヘミセルロースのうちマンナン系画分を標的にし、消化管内で高分子多糖が水を抱え込むことによる粘性上昇、栄養素拡散の低下、未消化多糖の後腸流入を緩和することです。Enzymes.bioの製品情報では、Mannanase Feed Additionをヘミセルロース処理、消化性支援、マンノースオリゴ糖生成に関連する飼料用酵素として位置づけています。
ここでいう「oligosaccharide processing」は、哺乳類細胞のゴルジ体で起こるN結合型糖鎖プロセシングではなく、飼料原料中のマンナン系多糖を酵素的に短鎖化する処理を指します。つまり、本品の実務的な焦点は、糖タンパク質修飾の制御ではなく、飼料マトリックス中の非デンプン性多糖を消化管環境で扱いやすい形へ変換することにあります。
β-マンナンが飼料中に存在すること自体は珍しくありません。問題は、その構造と物性が、動物の消化酵素だけでは十分に処理されにくい場合がある点です。単胃動物、とくに家禽や離乳後の豚では、可溶性または部分可溶性の非デンプン性多糖が消化管内容物の粘度を高め、消化酵素、胆汁酸、栄養素、腸上皮表面の接触を妨げることがあります。ヘミセルロース分解酵素は、このような植物細胞壁由来多糖を低分子化し、リグノセルロース系基質の利用性を高める技術群として研究されています[2]。
大豆粕はタンパク質源として広く使われますが、マンナン系多糖を含むため、配合比率や他の繊維性原料との組み合わせによっては、消化管内で抗栄養的な作用が顕在化します。パーム核粕やコプラミールのような副産物原料は、コスト・資源利用の観点で有用ですが、マンナンや関連ヘミセルロースが多く、単純に置換率を高めるとエネルギー利用や糞便性状に影響し得ます。パーム核粕をマンノースに富むシロップへ酵素的に変換する研究が行われていることは、この原料にマンナン系基質が豊富に含まれ、酵素分解の対象になり得ることを示しています[3]。
植物副産物を使う配合では、β-マンナンだけが唯一の制限要因ではありません。キシラン、セルロース、ペクチン、リグニン、タンパク質マトリックス、熱加工による構造変化が同時に関与します。したがってマンナナーゼの価値は、「すべての繊維を分解する」ことではなく、マンナン系ヘミセルロースという明確な基質に対して、粘性・包埋・未消化多糖流入の一部を下げる点にあります。
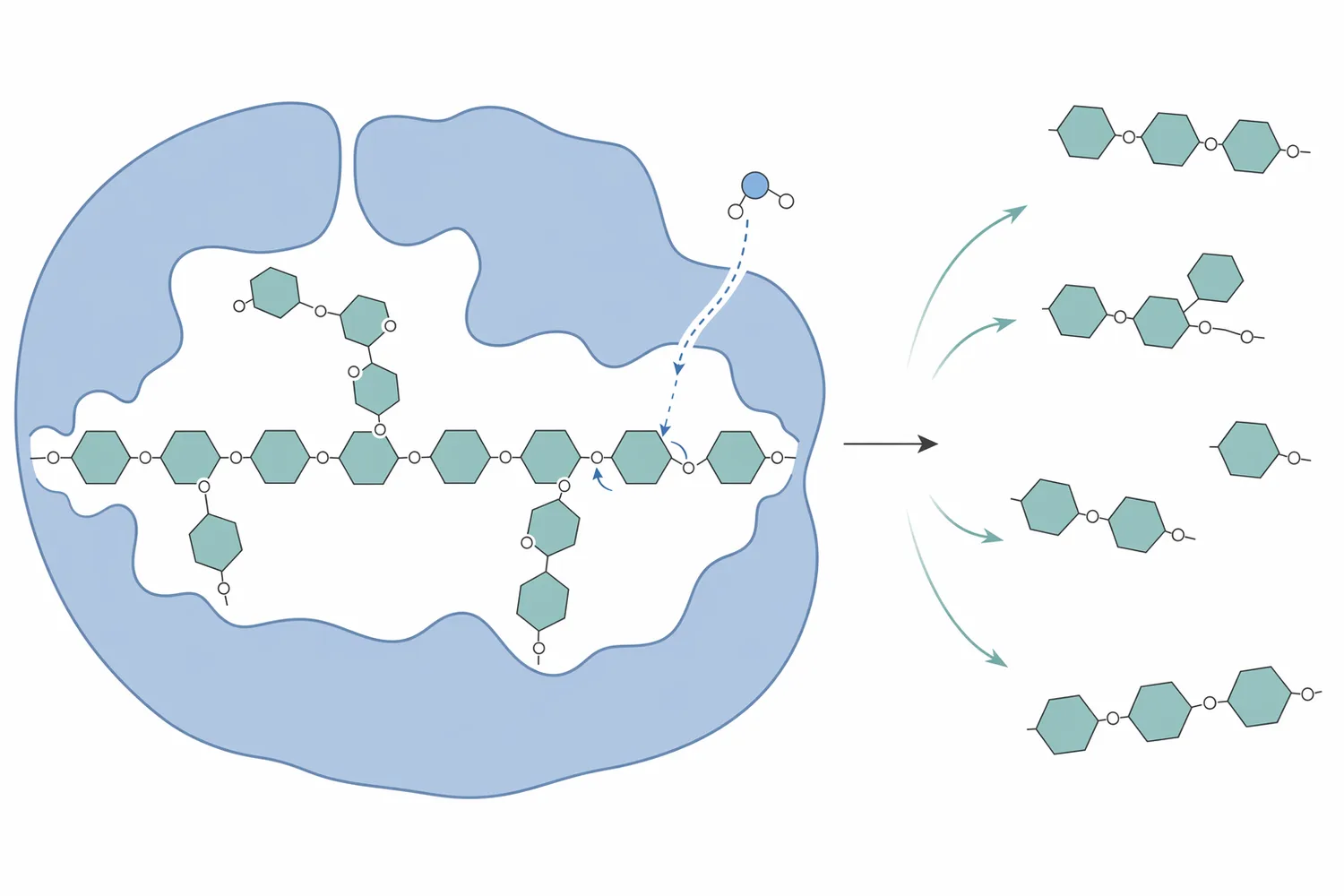
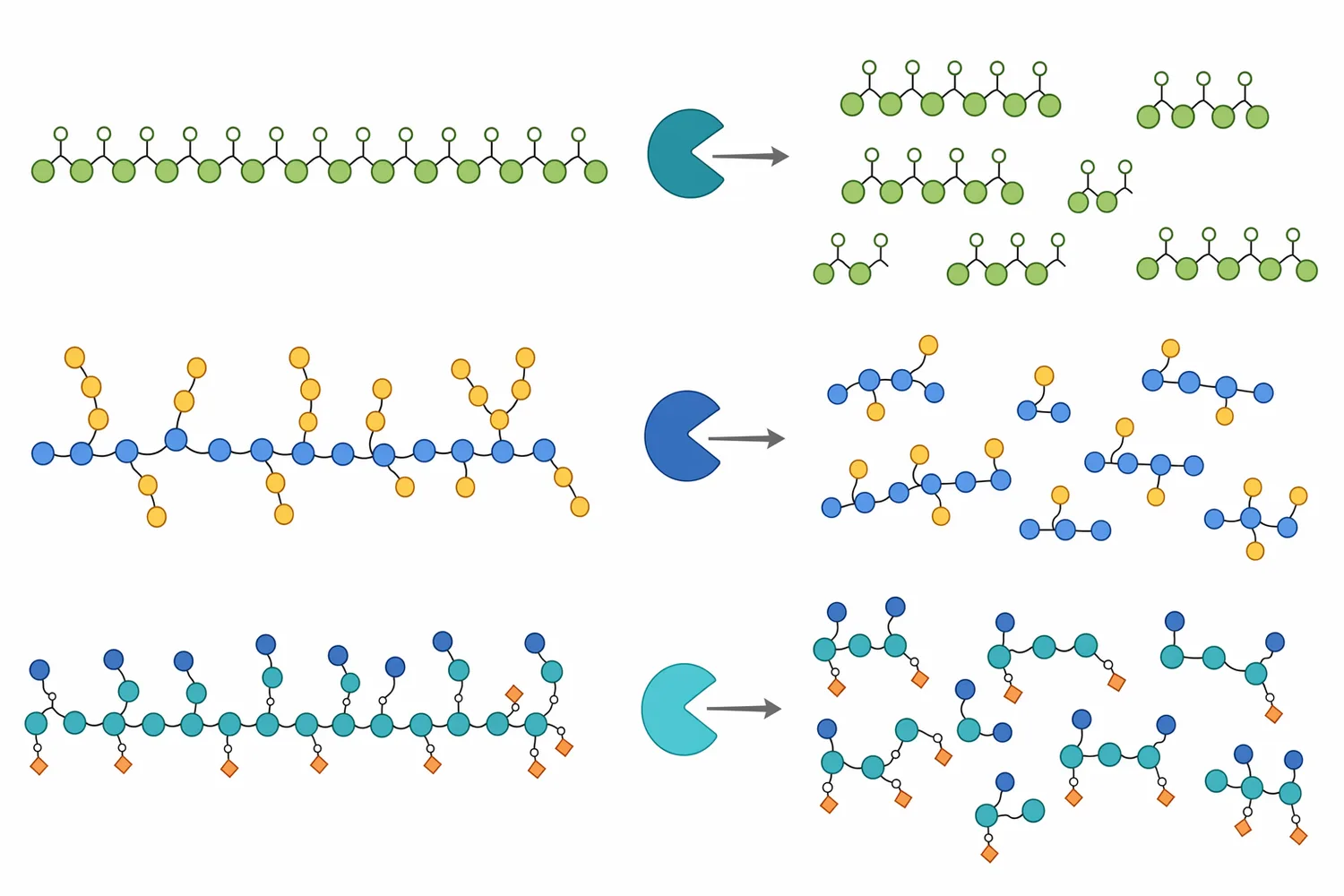
β-マンナナーゼの中心的な反応は、マンナン、ガラクトマンナン、グルコマンナンの主鎖にあるβ-1,4-マンノシド結合の加水分解です。多くの飼料用マンナナーゼはエンド型に分類され、糖鎖の末端から単糖を一つずつ外すのではなく、長い多糖鎖の内部結合を切断します。このため反応初期から分子量が急速に下がり、粘性の原因となる高分子鎖が短いマンノオリゴ糖へ移行します。endo-β-マンナナーゼの生産、性質、応用を扱うレビューでも、これらの酵素はマンナン系多糖の内部結合を標的にし、食品、飼料、バイオマス変換などで利用される酵素群として整理されています[4]。
粘性低減の機序は、単に「糖が増える」ことではありません。高分子多糖は長い鎖同士が絡み合い、水和層を形成し、消化管内容物の流動性を下げます。マンナナーゼが主鎖を複数箇所で切ると、鎖長が短くなり、分子間の絡み合いが減り、水を保持してゲル状または高粘度状態を作る能力が低下します。その結果、飼料粒子、内因性消化酵素、胆汁酸、腸上皮との接触が改善されやすくなります。
ただし、マンナンの分解しやすさは、主鎖だけで決まりません。ガラクトース側鎖が多いガラクトマンナンでは、β-マンナナーゼの基質結合溝への接近が妨げられる場合があります。パーム核粕の加水分解では、β-マンナナーゼとα-ガラクトシダーゼの相乗作用が報告されており、側鎖除去により主鎖切断が進みやすくなるという酵素設計上の考え方を示しています[5]。
マンナナーゼ処理によって生じる主な低分子生成物は、マンノースを含む短鎖糖、すなわちマンノオリゴ糖またはマンナン由来オリゴ糖です。飼料文脈では、これらが腸内微生物の発酵基質となり、プレバイオティック様に働く可能性が議論されます。ただし、生成物の鎖長、分岐、置換基、可溶性は原料ごとに異なり、同じ「マンノースオリゴ糖」と表現しても生理作用は一様ではありません。
重要なのは、マンノースオリゴ糖生成を過度に単純化しないことです。高分子マンナンが短鎖化されると粘性低減には寄与し得ますが、そのオリゴ糖がどの菌群に利用されるか、どの程度の短鎖脂肪酸産生につながるか、腸管免疫やバリア機能へどの程度影響するかは、動物種、日齢、腸内細菌叢、飼料全体の発酵性炭水化物量に依存します。ヒト腸内細菌叢由来酵素の解析でも、食品・バイオテクノロジー用途において糖質分解酵素が基質構造に依存して多様な生成物を生むことが示されており、オリゴ糖の機能は構造と微生物利用性を切り離して評価できません[6]。
したがって、本品の主要な説明軸は「β-マンナン系ヘミセルロースの低分子化による消化環境の支援」であり、マンノースオリゴ糖生成はその結果として期待される二次的価値です。プレバイオティック様作用を想定する場合でも、配合設計や飼養条件により反応が変動するため、万能な腸内改善効果として表現するのではなく、マンナン基質が十分に存在する条件で成立しやすい可能性として扱うのが妥当です。
飼料中のマンナンは、原料の種類によって存在形態が異なります。大豆粕では、タンパク質源としての価値が高い一方、細胞壁多糖が消化性の上限を作ることがあります。パーム核粕やコプラミールでは、マンナン系多糖の寄与が大きく、エネルギー利用性を制限しやすい一方で、適切に酵素処理されれば植物副産物の利用範囲を広げる余地があります。農産副産物を飼料へ活用する近年の研究では、酵素支援によって繊維組成や消化性を改善しようとする方向性が複数の動物種で検討されています[7]。
| 飼料原料・配合場面 | マンナン関連の課題 | マンナナーゼで期待される主な作用 | 注意すべき点 |
|---|---|---|---|
| 大豆粕主体の家禽・豚用配合 | β-マンナンによる粘性、栄養素拡散の低下、抗栄養的影響 | マンナン主鎖の低分子化、消化管内容物の流動性支援 | 大豆粕品質、加熱履歴、他の非デンプン性多糖の影響も受ける |
| パーム核粕を含む配合 | マンナン系ヘミセルロースが多く、エネルギー利用が制限されやすい | マンノオリゴ糖・低分子糖鎖への変換、繊維性副産物の利用支援 | 側鎖構造や粒度により酵素アクセス性が変わる |
| コプラミール・繊維性副産物 | 高繊維、複合細胞壁、可消化エネルギーのばらつき | マンナン画分の分解による消化負荷の低減 | キシラナーゼ、セルラーゼ、ペクチナーゼ等との役割分担が必要な場合がある |
| 植物副産物を多く使うサステナブル配合 | 原料コスト低減と消化性低下のトレードオフ | ヘミセルロース処理による配合自由度の支援 | 問題成分がマンナン以外なら単独効果は限定的 |
トウモロコシ外皮やコーンファイバーのような材料では、ヘミセルロース側鎖の構造が酵素カクテル設計を左右することが報告されています。これは、マンナナーゼの選択が単なる「繊維分解酵素の追加」ではなく、原料中の糖鎖構造と酵素特異性を対応させる作業であることを示しています[8]。
家禽では、飼料摂取量、腸管通過速度、消化管内容物の粘性が成績に直結しやすいため、非デンプン性多糖の管理が重要です。β-マンナンが多い配合では、内容物粘度の上昇により消化酵素と基質の接触が悪くなり、エネルギー利用が下がる可能性があります。家禽向けの産業教育資料でも、β-マンナナーゼは植物性飼料に含まれるβ-マンナンの抗栄養的影響を下げ、腸管環境と飼料効率を支援する酵素として説明されています[9]。
近年、ブロイラー生産では、農産副産物の利用、飼料コスト、環境負荷を同時に考える必要が高まっています。酵素支援による副産物の価値化を扱う研究では、植物性副産物の繊維画分を酵素的に扱うことで、持続的かつ効率的なブロイラー生産へつなげる方向性が示されています[10]。
ただし、家禽でマンナナーゼを使う意味は、すべての配合で一律の成績改善を約束することではありません。マンナン含量が低い配合、消化性の高い精製原料を多く使う配合、あるいは疾病・温熱ストレス・水質などが主因の不調では、マンナナーゼの寄与は見えにくくなります。効果を期待しやすいのは、マンナンを含む植物性タンパク源や副産物原料が、消化管粘性や栄養利用の制限要因になっている場合です。
豚では、とくに離乳後に消化管機能と腸内細菌叢が不安定になりやすく、未消化の非デンプン性多糖が後腸へ流入すると、発酵パターンや糞便性状に影響します。β-マンナナーゼを配合する目的は、マンナン系ヘミセルロースを前段階で低分子化し、内容物粘性や栄養素包埋を緩和することです。離乳後子豚におけるβ-マンナナーゼ添加の研究では、ネットエネルギーを下げた飼料でも同等の成績と経済的利点が報告されており、マンナン分解がエネルギー設計に関与し得ることを示しています[11]。
スターター豚の飼料では、商業飼料の一部を農産副産物ベースの原料で置き換える試みもあります。副産物利用では、コストや資源循環の利点と、繊維性成分による消化性低下が同時に存在します。農産副産物ベース飼料の最適化研究は、こうした代替原料を単純な置換ではなく、栄養価、繊維、酵素利用を含む設計課題として扱う必要があることを示しています[12]。
豚用配合でのマンナナーゼは、タンパク質消化酵素や有機酸、プロバイオティクスと同じ機序で働くものではありません。標的はあくまでマンナン系多糖であり、効果は原料のβ-マンナン量、加熱処理、粉砕粒度、他の酵素との組み合わせに依存します。そのため、マンナナーゼの実務的価値は、離乳後の脆弱な消化環境において、特定の植物性ヘミセルロース負荷を下げることにあります。
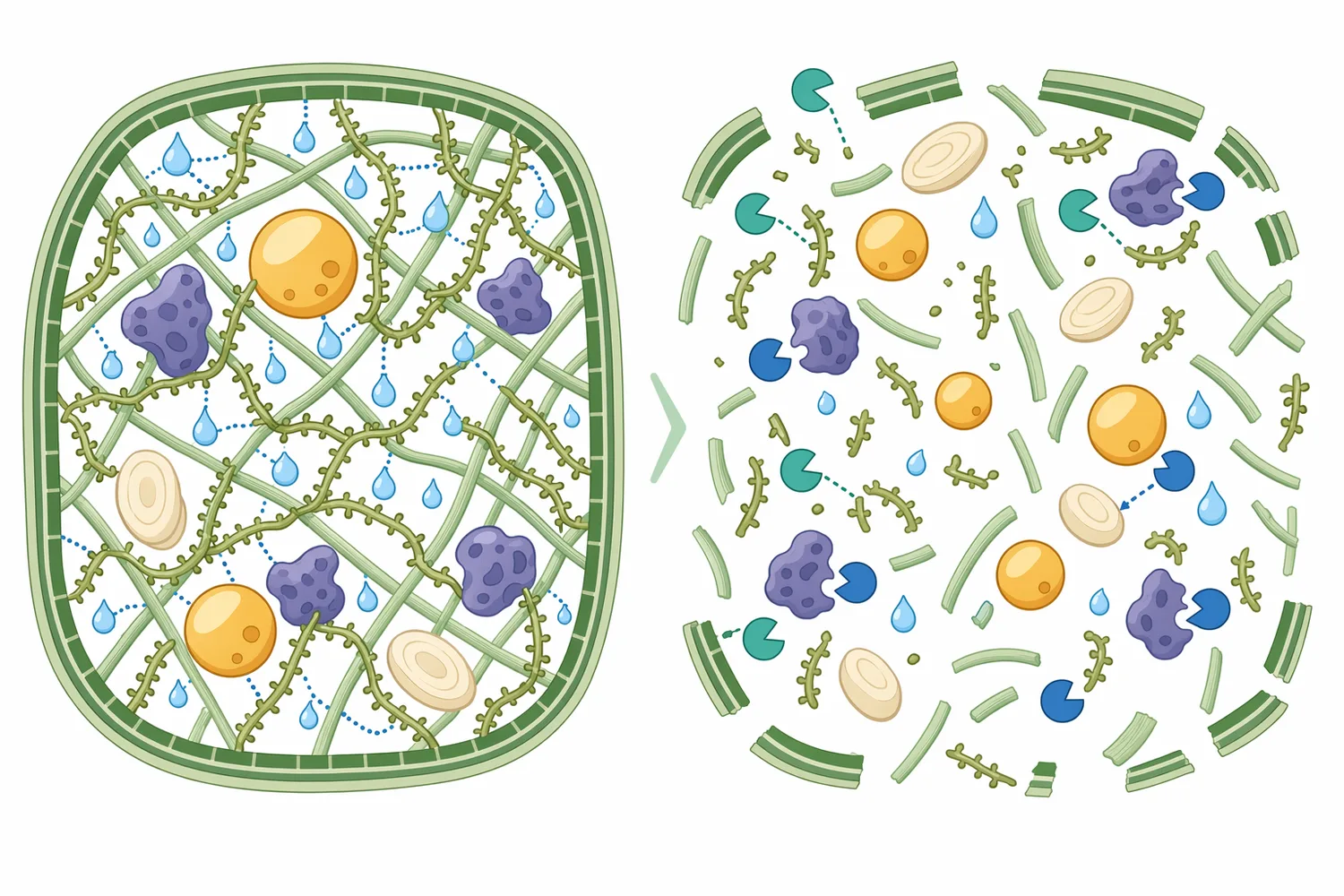
植物細胞壁は、マンナンだけで構成されているわけではありません。キシラン、セルロース、ペクチン、アラビノガラクタン、リグニンとの複合体として存在し、どの酵素が最も重要かは原料によって変わります。ヘミセルロース分解酵素に関するレビューでは、リグノセルロース処理において複数の酵素が基質の異なる結合や側鎖を処理し、全体の分解効率を左右することが説明されています[2]。
マンナナーゼは、キシラナーゼやセルラーゼの代替ではありません。キシランが主な制限要因であればキシラナーゼが中心となり、セルロースの露出が問題であればセルラーゼや前処理条件が重要になります。マンナナーゼが最も意味を持つのは、マンナン、ガラクトマンナン、グルコマンナンが粘性や栄養素包埋の主要因になっている配合です。コーンファイバー由来の難分解性ヘミセルロースオリゴ糖を扱う研究でも、構造解析に基づいて酵素戦略を組み立てる必要性が示されています[13]。
| 酵素 | 主な標的 | マンナナーゼとの関係 | 飼料設計上の見方 |
|---|---|---|---|
| β-マンナナーゼ | マンナン、ガラクトマンナン、グルコマンナン主鎖 | 本品の中心機能 | 大豆粕、パーム核粕、コプラミールなどで重要 |
| α-ガラクトシダーゼ | ガラクトース側鎖 | ガラクトマンナン分解を助ける場合がある | 側鎖が多い基質で相乗性が期待される |
| キシラナーゼ | アラビノキシラン、キシラン | 標的が異なる補完酵素 | 小麦、ライ麦、トウモロコシ副産物などで検討される |
| β-グルカナーゼ | β-グルカン | 粘性多糖対策として補完的 | 大麦、オート麦系配合で重要になることがある |
| セルラーゼ | セルロース | 細胞壁開放に関与し得る | 高繊維副産物では補助的に扱われる |
この役割分担を理解すると、マンナナーゼを「繊維分解酵素」と一括りにするよりも、どの多糖構造に対して効果を狙うのかが明確になります。酵素工学の産業応用では、基質特異性、安定性、反応条件を目的用途に合わせて最適化する考え方が重視されており、飼料酵素でも同じく、酵素名だけでなく対象基質と使用環境の一致が重要です[14]。
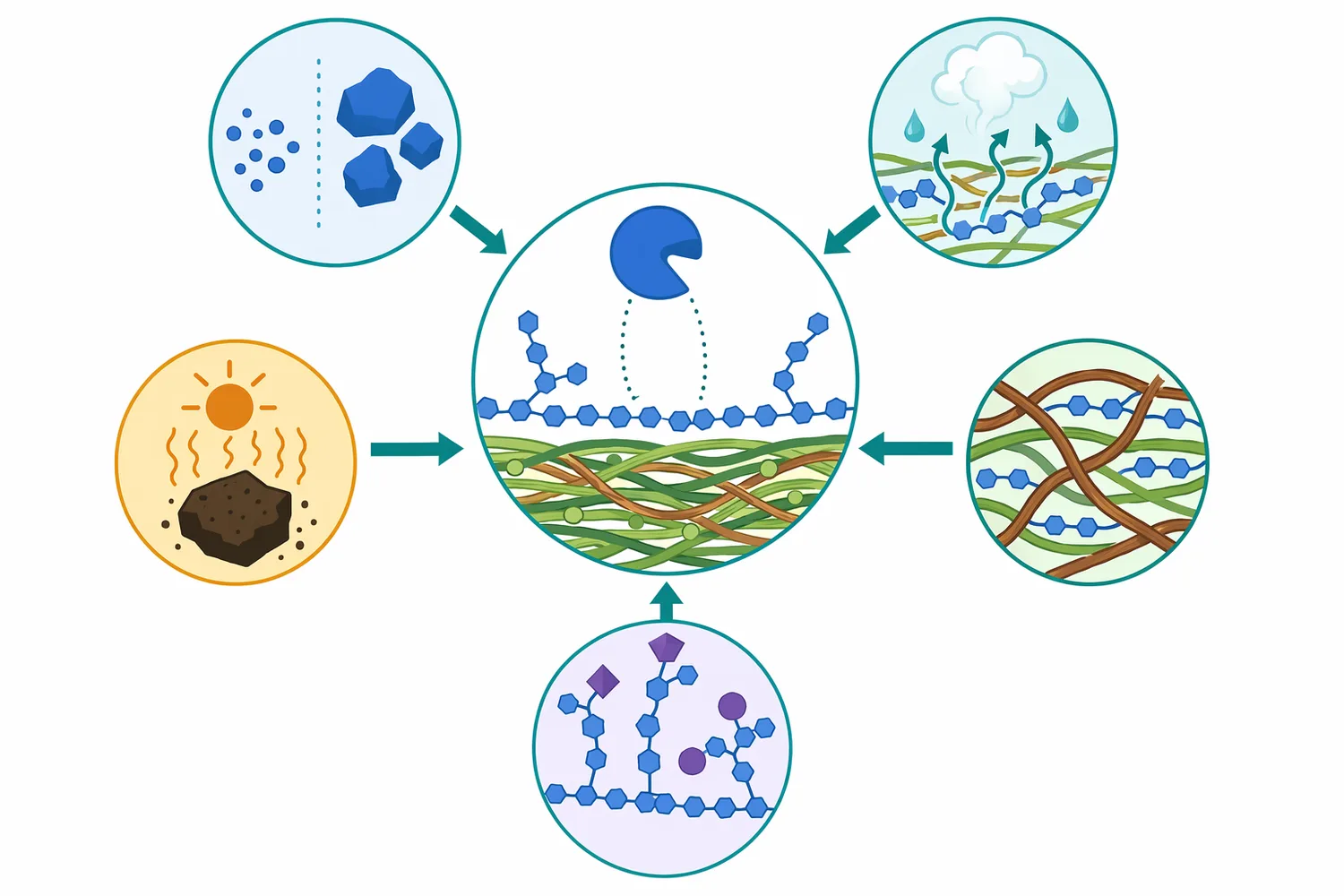
マンナナーゼの反応は、基質の存在量だけでなく、飼料加工と消化管環境の影響を受けます。ペレット化、エクストルージョン、加熱乾燥、原料の水熱処理は、ヘミセルロースの溶出、再凝集、側鎖の変化、細胞壁マトリックスの開放に関与します。水熱処理がヘミセルロース構造へ影響することは、バイオマス処理研究でも詳細に扱われており、加工条件によって酵素がアクセスできる糖鎖構造が変わることを示しています[15]。
消化管内では、pH、滞留時間、水分、胆汁、内因性酵素、微生物発酵が同時に作用します。マンナナーゼが十分に機能するには、飼料摂取後の環境で酵素構造が保たれ、基質と接触できる必要があります。endo-β-マンナナーゼのpH依存的安定性を扱う研究では、酵素の構造安定性が環境条件に左右されることが示されており、飼料用途でも「酵素が存在すること」と「消化管内で基質を処理できること」は同義ではありません[16]。
同時に、熱安定性やpH安定性だけで効果は決まりません。酵素が耐性を持っていても、マンナンがリグニンやタンパク質マトリックスに包埋されていれば反応は制限されます。逆に、加工により細胞壁が開放され、適度に水和した基質では、酵素とマンナン鎖の接触が増えます。したがって、実際の効果は酵素特性、基質構造、加工履歴、動物側の消化環境が重なった結果として理解する必要があります。
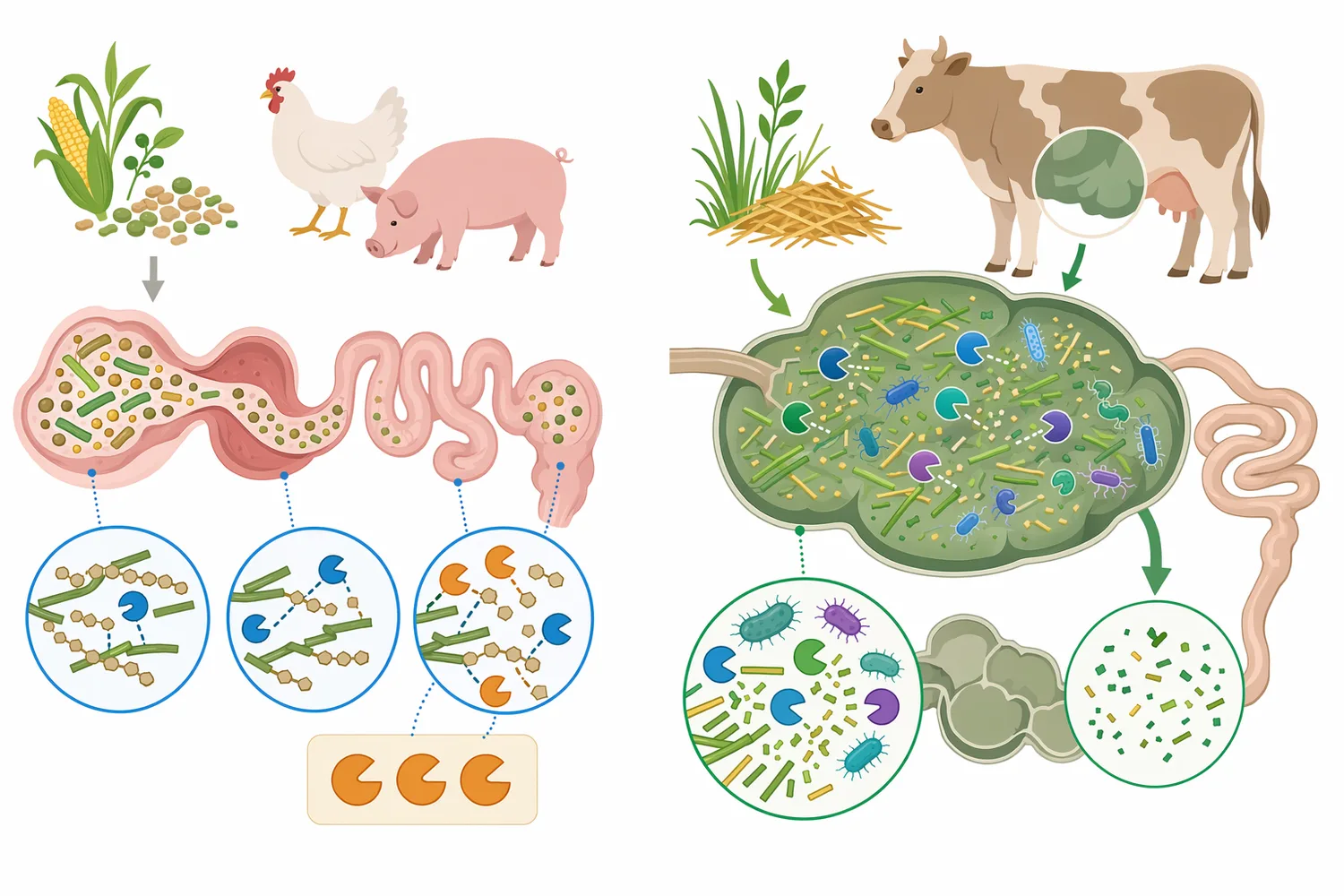
マンナナーゼに関する根拠は、いくつかの層に分けて読むと整理しやすくなります。第一に、酵素学的根拠として、β-マンナナーゼがβ-1,4-マンナン主鎖を加水分解することは確立された機能です。第二に、基質根拠として、大豆粕、パーム核粕、コプラミールなどにマンナン系ヘミセルロースが存在し、消化性を制限し得ることがあります。第三に、飼料成績根拠として、動物試験で飼料効率、エネルギー利用、糞便性状などに影響が見られる場合がありますが、これは配合と飼養条件に大きく依存します[4]。
特に注意すべきは、バイオマス糖化研究と飼料試験を混同しないことです。バイオマス研究は、マンナンやヘミセルロースが酵素で分解できることを強く支持しますが、そのまま動物成績の数値を保証するものではありません。一方、動物試験は飼料としての有用性を示しますが、原料組成や飼養条件が変わると結果も変わります。酵素支援による農産副産物利用を扱う研究が増えていることは、飼料コストと消化性の両立が現実的な課題であることを示しています[10]。
本品の説明では、こうした根拠を「マンナンを分解できる」「マンナン系原料で消化負荷を下げ得る」「条件により飼料利用を支援し得る」という順序で捉えるのが適切です。過度に単純な成績保証ではなく、基質特異性に基づく飼料酵素として理解することで、マンナナーゼの強みと限界が明確になります。
マンナナーゼは、飼料中のすべての抗栄養因子を取り除く酵素ではありません。タンパク質の熱損傷、脂質酸化、マイコトキシン、ミネラル不均衡、病原体圧、飼養管理の問題は、マンナン分解とは別の課題です。飼料成績の低下がこれらに由来する場合、マンナナーゼの添加だけで問題が解決するとは考えるべきではありません。
また、マンナン含量が少ない配合では、効果は小さくなり得ます。マンナナーゼは基質があって初めて働くため、精製度の高い原料や低繊維配合では、期待される粘性低減やオリゴ糖生成の余地が限られます。逆に、パーム核粕、コプラミール、特定の大豆副産物など、マンナン系ヘミセルロースの寄与が大きい原料では、酵素反応の意味が明確になります。
マンノースオリゴ糖生成についても、生成量や鎖長を一律に想定することはできません。原料の側鎖構造、加工条件、消化管内での滞留時間が変われば、同じマンナナーゼでも生成物プロファイルは変わります。したがって、本品は「マンノースオリゴ糖を必ず一定量作る添加物」ではなく、マンナン系多糖を低分子化し、その過程でマンノオリゴ糖生成を支援し得る酵素として表現するのが科学的です。
Enzymes.bioは、本品をB2B向けにオンライン供給する酵素サプライヤーであり、製造業者または試験研究機関として本稿を提示しているわけではありません。Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processingは、製品ページから1 kg単位で直接購入できる飼料用途向け酵素として掲載されています。
注文時には、CoAおよびSDSが併せて提供されます。これらは受領後の社内記録、保管、取り扱い安全情報の管理に用いられる文書であり、本稿では個別ロットの数値、分析法、活性単位定義、製造条件を記載しません。Enzymes.bioのマンナナーゼ関連カテゴリでは、マンナナーゼが飼料、食品加工、植物原料処理など複数の産業用途で扱われる酵素群として整理されています。
本品を位置づけるうえで重要なのは、供給形態と技術的用途を分けて理解することです。供給形態としてはオンラインで1 kg単位の直接販売、技術的用途としては飼料中のβ-マンナン系ヘミセルロースの低分子化です。製造者のように菌株開発、発酵製造、個別分析手法を説明するのではなく、飼料設計でどの基質に作用し、どのような消化上の負荷を下げるかを明確にすることが、この製品情報の目的です。
Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processingは、飼料中のマンナン、ガラクトマンナン、グルコマンナンを低分子化し、消化管内容物の粘性負荷低減、栄養素利用の支援、マンナン由来オリゴ糖生成を狙う飼料用マンナナーゼです。β-マンナナーゼがマンナン系多糖を加水分解することは酵素学的に確立しており、ヘミセルロース分解酵素としてリグノセルロース処理や飼料原料活用の文脈で研究されています[2]。
実務上の価値が最も出やすいのは、大豆粕、パーム核粕、コプラミール、繊維性農産副産物など、マンナン系ヘミセルロースが消化性の制限要因になり得る配合です。家禽や豚では、マンナンによる粘性、栄養素包埋、未消化多糖の流入を下げる補助的手段として利用され、離乳後子豚などではエネルギー設計と関連する研究も報告されています[11]。
一方で、マンナナーゼは万能な成績改善剤ではありません。効果は基質量、原料構造、加工履歴、動物種、消化管環境、他酵素との組み合わせに依存します。そのため、本品は「特定のマンナン系ヘミセルロースに作用する専門酵素」として扱うのが正確であり、植物性原料を含む飼料の利用性を支援するための現実的な酵素ソリューションとして位置づけられます。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processingを購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。