직접 답변: Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processing은 식물성 원료의 만난계 헤미셀룰로오스, 특히 β-1,4-만난 골격을 더 짧은 만노올리고당과 일부 만노스 계열 산물로 절단하는 β-만나나아제 기반 효소 제품으로 이해할 수 있습니다. 사료에서는 대두박, 구아 meal, soy hulls, 팜계 부산물처럼 만난성 비전분다당류가 소화율·점도·영양소 손실에 영향을 줄 수 있는 원료에서 검토되며, 바이오매스 처리에서는 만난 풍부 기질을 올리고당 또는 발효성 당 전구체로 전환하는 공정에 연결됩니다 [1] [2].
Enzymes.bio는 이 제품의 공급업체이며 제조사나 분석 실험실이 아닙니다. 제품은 온라인에서 1kg 단위로 직접 구매할 수 있고, 주문 시 CoA와 SDS가 함께 제공됩니다 .
식물성 사료와 농산 부산물의 세포벽은 셀룰로오스 미세섬유, 헤미셀룰로오스, 펙틴, 리그닌이 서로 얽힌 복합 구조입니다. 이 중 만난, 갈락토만난, 글루코만난, 갈락토글루코만난은 헤미셀룰로오스의 한 축을 이루며, 만노스 잔기가 β-1,4 결합으로 연결된 주사슬을 갖습니다. 식물의 2차 세포벽은 조직 강도와 방어 기능을 위해 두껍게 축적되며, 이 구조가 사료 소화와 효소 접근성을 동시에 제한할 수 있습니다 [3].
β-만나나아제는 이 만난 주사슬 내부의 β-1,4-만노시드 결합을 절단하는 endo형 가수분해효소로 설명됩니다. 즉, 사슬 끝에서 한 단위씩 떼어내는 방식이 아니라 고분자 사슬 중간을 여러 지점에서 끊어 분자량과 중합도를 낮춥니다. 그 결과 긴 만난 고분자는 더 짧은 만노올리고당, 경우에 따라 일부 만노스 또는 더 작은 당질 조각으로 바뀌며, 완전한 단당화를 위해서는 β-만노시다아제 같은 보조 효소가 추가로 필요할 수 있습니다 [4].
이 효소의 실무적 의미는 “섬유를 없앤다”가 아니라 “만난성 헤미셀룰로오스의 물리·영양학적 영향을 바꾼다”에 가깝습니다. 만난계 다당류가 물을 붙잡고 점성을 높이거나, 식물성 세포벽 내부에 영양소를 가두거나, 장내에서 비전분다당류 부담을 만드는 경우 β-만나나아제는 그 구조를 짧게 잘라 영향력을 낮출 수 있습니다. 리그노셀룰로오스 분해 연구에서도 세포벽 해체는 단일 반응이 아니라 효소 위치, 기질 접근성, 벽 성분의 화학적 변화가 함께 작용하는 과정으로 다뤄집니다 [5].
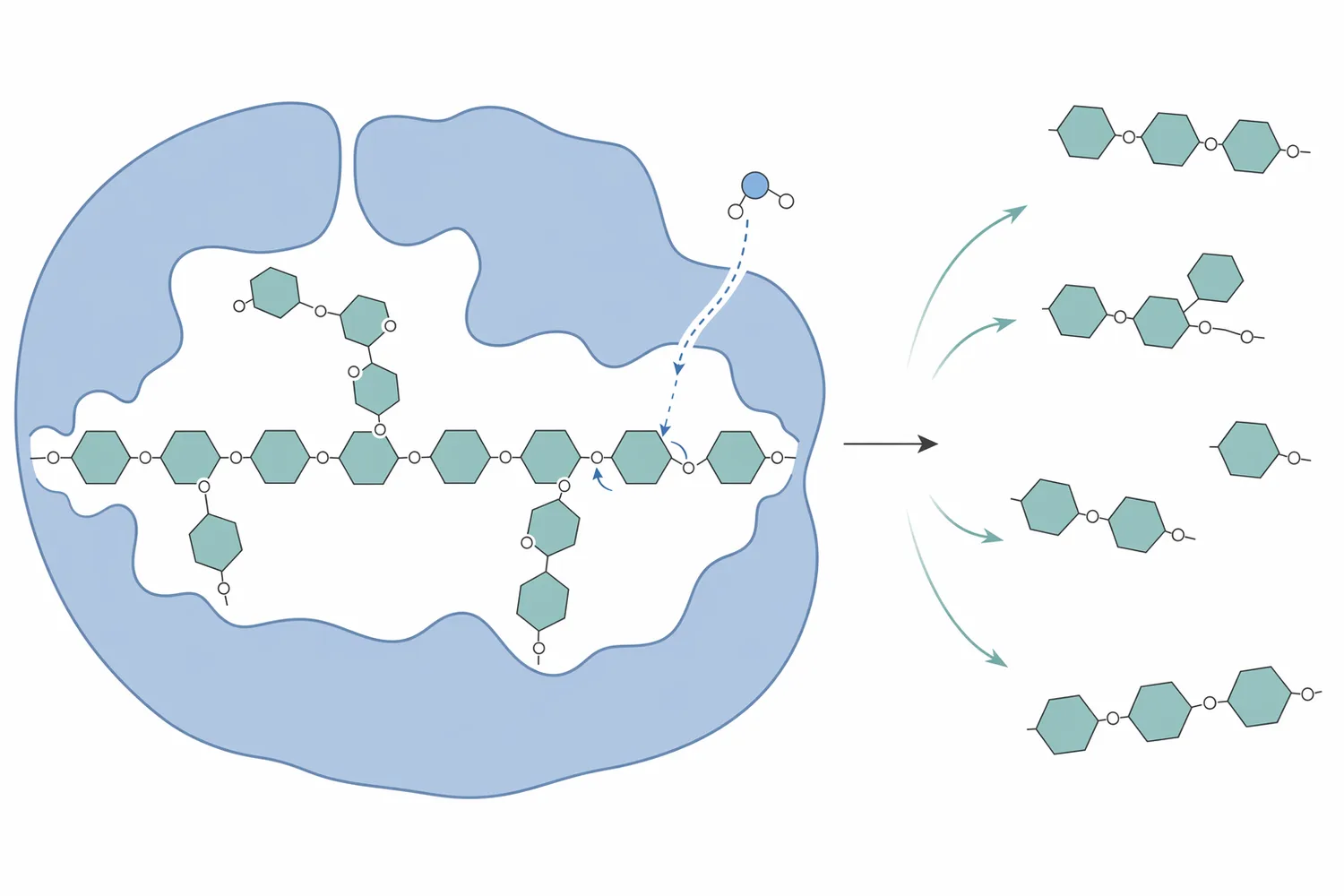
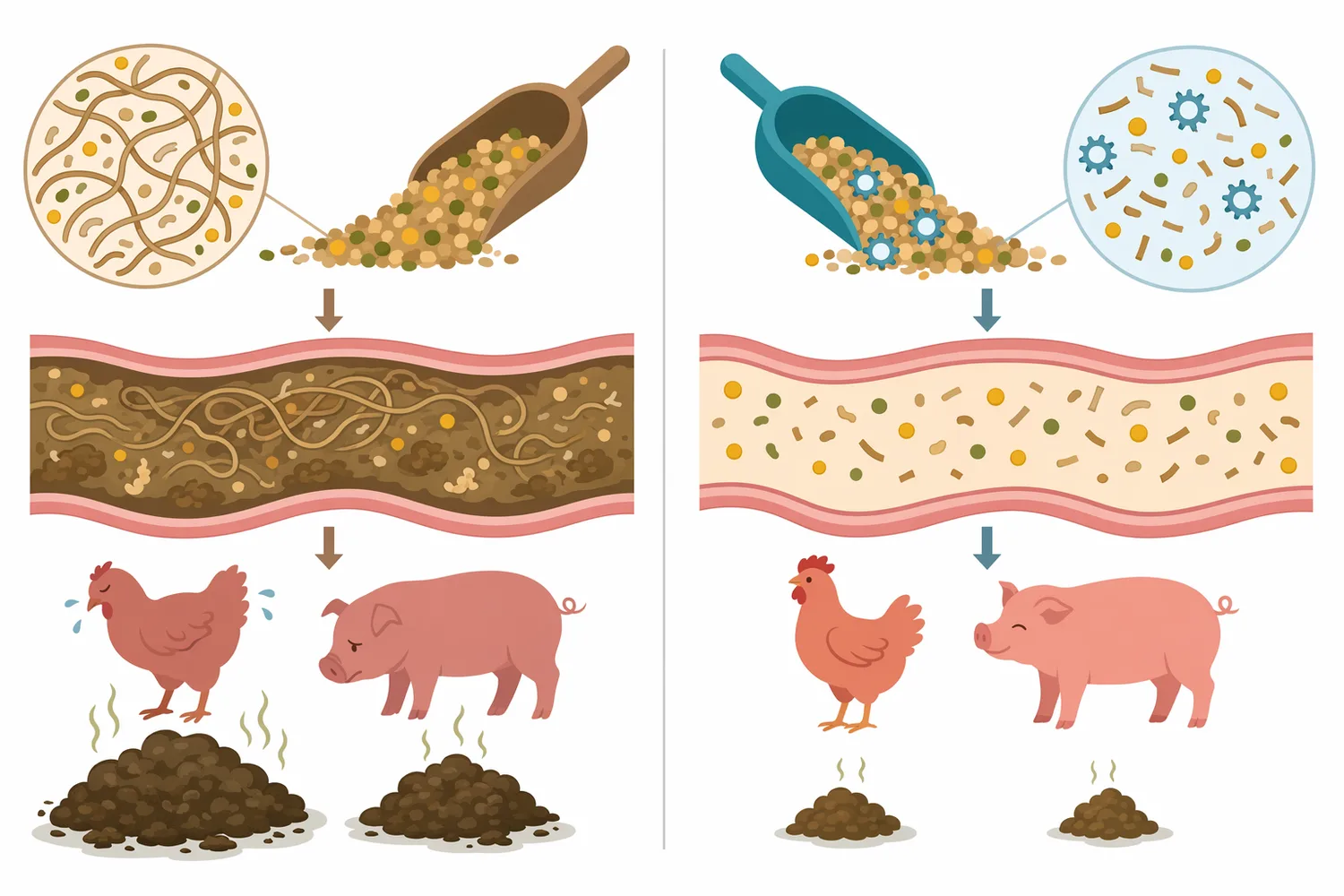
식물성 사료 원료의 조단백, 조지방, 에너지 값이 충분해 보여도 실제 동물의 이용률은 비전분다당류 구조에 의해 달라질 수 있습니다. β-만난은 대두 유래 원료, 구아 meal, soy hulls, 팜계 부산물 등 여러 식물성 원료에서 문제 될 수 있으며, 단위동물은 이런 구조성 다당류를 자체 효소만으로 충분히 분해하지 못합니다. 돼지 사료 연구에서는 phytase와 β-mannanase 보충이 성장 성능, 영양소 이용, 분변 상태, 등지방 두께와 함께 평가되었고, 이는 β-만난 저감이 실제 배합 사료 설계에서 검토되는 항목임을 보여줍니다 [1].
가금류에서도 유사한 논리가 적용됩니다. 육계는 고섬유 또는 특정 비전분다당류가 높은 사료를 섭취할 때 장내 내용물의 점성, 영양소 확산, 소화효소와 기질의 접촉 효율에 영향을 받을 수 있습니다. Soy hulls 수준과 β-mannanase 보충을 함께 평가한 육계 연구는 성장 성능, 도체 특성, 영양소 이용, 혈액 생화학 지표를 함께 관찰했으며, 이는 β-만나나아제가 단순한 “섬유 분해제”가 아니라 식물성 원료 사용 폭과 영양 이용성을 조정하는 도구로 연구되고 있음을 시사합니다 [6].
구아 meal은 갈락토만난 성격이 강한 원료로, 만난계 점질 성분과 관련된 사료 가공성·소화성 문제가 특히 두드러질 수 있습니다. 구아 meal을 섭취한 육계에서 dietary enzyme 및 fermentation treatment가 성능, 소화율, 생화학 프로필에 미치는 영향을 평가한 연구는, 만난성 원료가 들어간 배합에서 효소 처리와 발효 처리가 함께 검토될 수 있음을 보여줍니다 [7].
수산사료에서도 β-만난 문제는 무관하지 않습니다. 대두박 비중이 높은 juvenile Nile tilapia 사료에서 β-mannanase가 분변 점도, 소화에너지, 아미노산 소화율, 영양소 손실에 미치는 영향을 다룬 연구는, 만난계 비전분다당류가 육상 단위동물뿐 아니라 어류의 식물성 단백질 원료 활용에서도 고려될 수 있음을 보여줍니다 [2].
만난계 헤미셀룰로오스의 핵심은 β-1,4로 연결된 만노스 기반 주사슬입니다. β-만나나아제가 이 주사슬 내부를 절단하면, 같은 질량의 다당류라도 평균 사슬 길이가 줄어듭니다. 긴 사슬이 짧아지면 용액 내 얽힘과 수화 네트워크가 약해지고, 갈락토만난처럼 점질성을 띠는 기질에서는 흐름성과 혼합성이 달라질 수 있습니다. 식물 세포벽 효소 가수분해 연구에서는 이러한 분자 수준의 절단이 세포벽 구조 변화와 기질 접근성 변화로 이어진다고 설명합니다 [4].
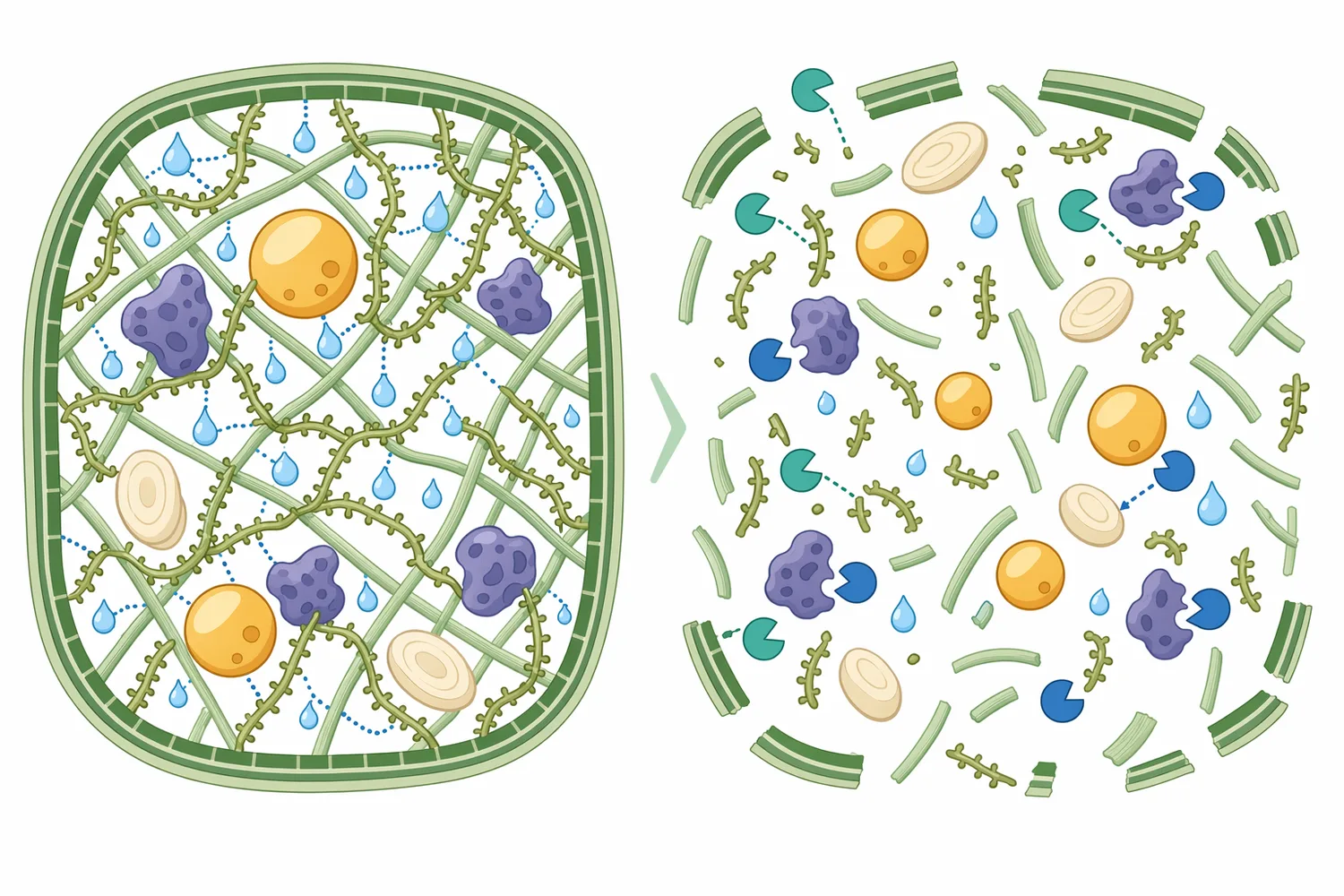
사료 원료의 영양소는 세포벽 안팎에 물리적으로 갇혀 있을 수 있습니다. 셀룰로오스 미세섬유는 구조적 골격을 제공하고, 헤미셀룰로오스와 펙틴은 그 사이를 메우며, 리그닌과 페놀성 교차결합은 효소 접근을 더 어렵게 만듭니다. 식물 세포벽 내 diferulate 같은 교차결합 물질은 벽 성분 간 연결성을 높이는 요소로 분석되어 왔으며, 이런 복합성 때문에 한 가지 효소만으로 전체 세포벽을 완전히 해체하기는 어렵습니다 [8].
β-만나나아제의 역할은 이 복합 매트릭스 중 만난계 연결을 선택적으로 약화시키는 것입니다. 만난이 영양소 주변의 점질 장벽 또는 구조적 포획 요소로 작용할 때, 만난 사슬 절단은 소화효소·미생물·수분이 기질에 접근하는 방식을 바꿀 수 있습니다. 다만 원료의 주된 제한 요인이 자일란이면 자일라아제, 셀룰로오스 결정성이라면 셀룰라아제, 피틴산이라면 피타아제가 더 직접적일 수 있으므로 β-만나나아제는 “만난 표적 효소”로 해석하는 것이 정확합니다 [9].
β-만나나아제 처리의 산물은 흔히 만노올리고당, 즉 mannooligosaccharides 또는 MOS로 설명됩니다. 여기서 MOS는 효소가 식물성 만난을 절단해 만든 올리고당 산물을 의미하며, 효모 세포벽 유래 MOS나 세포 내 N-linked oligosaccharide processing과는 맥락이 다릅니다. 골지체에서의 N-linked oligosaccharide processing은 당단백질 성숙과 표적화에 관한 세포생물학적 과정으로 연구되어 왔지만, 본 제품명에서 말하는 mannose oligosaccharide processing은 사료·바이오매스 기질의 만난계 다당류 처리에 가깝습니다 [10].
만노올리고당은 그 자체로 사료·발효·기능성 당질 소재 연구의 중간산물이 될 수 있고, 추가 효소나 미생물 공정을 거쳐 더 작은 당으로 전환될 수도 있습니다. 하지만 β-만나나아제 단독 처리만으로 모든 만난이 만노스 단당으로 완전히 바뀐다고 보는 것은 과도합니다. 목표가 점도 완화인지, MOS 생성인지, 발효성 당 생산인지에 따라 필요한 효소 조합과 공정 조건은 달라집니다 [11].
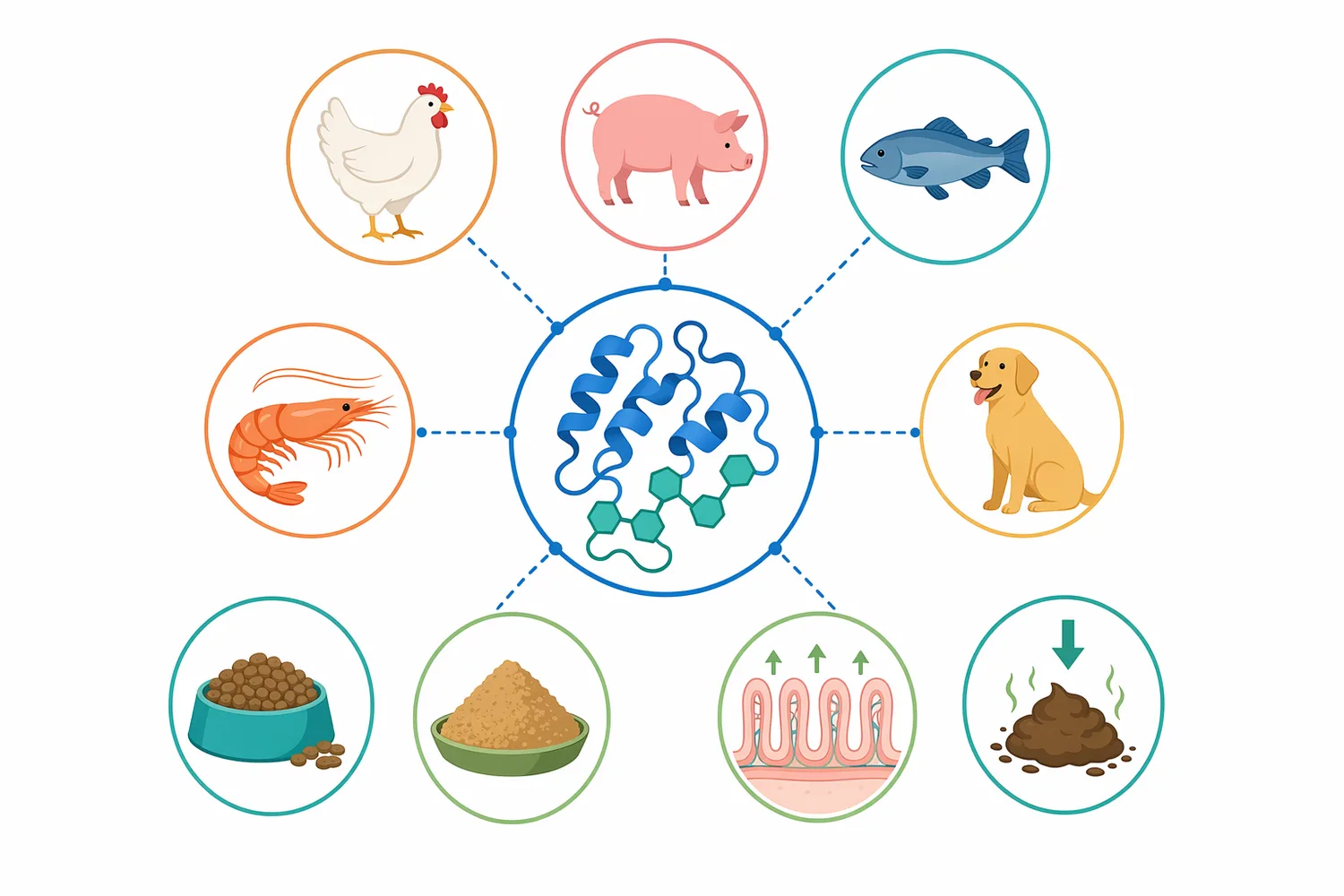
| 적용 분야 | 주된 기질 또는 원료 | β-만나나아제의 역할 | 기대되는 기술적 변화 | 해석 시 주의점 |
|---|---|---|---|---|
| 돼지 사료 | 식물성 원료, 만난 함유 부산물 | β-만난 절단 | 영양소 이용, 분변 상태, 성장 지표와 함께 평가 | 원료 조성·성장 단계·배합 에너지에 따라 결과 달라짐 [1] |
| 육계 사료 | soy hulls, 구아 meal 등 | 만난계 NSP 부담 완화 | 성장 성능, 소화율, 혈액 지표, 도체 특성과 연계 평가 | 고섬유 원료의 전체 제한 요인이 만난인지 확인 필요 [7] |
| 수산사료 | 대두박 비중이 높은 어류 사료 | 점도·영양소 손실 관련 만난 저감 | 분변 점도, 소화에너지, 아미노산 소화율과 연계 | 어종별 장 생리와 식물성 단백질 적응성 차이 큼 [2] |
| 헤미셀룰로오스 처리 | 만난, 갈락토만난, 글루코만난 | β-1,4-만난 골격 가수분해 | 분자량 감소, MOS 생성, 접근성 변화 | 리그닌·셀룰로오스·자일란 분해는 별도 효소 필요 [4] |
| 바이오매스 전환 | 농산 부산물, 섬유성 원료 | 만난 분획의 당화 보조 | 올리고당 또는 발효성 당 전구체 형성 | 전체 리그노셀룰로오스 전환에는 복합 효소 전략 필요 [5] |
이 표의 핵심은 β-만나나아제가 “전체 섬유 분해제”가 아니라 “만난계 헤미셀룰로오스의 특정 결합을 끊는 효소”라는 점입니다. 따라서 적용 판단은 원료명만으로 하기보다 그 원료에서 만난성 NSP가 실제 제한 요인으로 작용하는지에 따라 달라집니다. 예를 들어 soy hulls는 섬유성 원료이지만 모든 섬유가 만난인 것은 아니며, 구아 meal은 갈락토만난 관련성이 더 직접적일 수 있습니다 [6].
대두박은 단백질 공급원으로 널리 사용되지만, 단백질만 보는 배합에서는 비전분다당류의 물리적 영향이 과소평가될 수 있습니다. 대두박이 풍부한 틸라피아 사료에서 β-mannanase가 분변 점도와 소화에너지, 아미노산 소화율, 영양소 손실에 미치는 영향을 평가했다는 점은, 대두 유래 원료의 만난성 성분이 단백질 이용성 논의와 분리될 수 없음을 보여줍니다 [2].
β-만나나아제는 대두박 자체를 “고단백 원료”에서 “더 이용 가능한 고단백 원료”로 바꾼다고 단정할 수는 없습니다. 그러나 만난계 NSP가 장내 점도와 영양소 확산을 방해하거나, 소화되지 않는 섬유성 분획으로 배출되는 경우에는 해당 분획을 짧게 절단해 소화 환경을 바꾸는 방향으로 작용할 수 있습니다. 이때 관찰 지표는 성장률 하나가 아니라 에너지 이용, 아미노산 소화율, 분변 또는 장 내용물 특성까지 포함해야 합니다 [2].
구아 meal은 만난계 점질 성분이 실무적으로 문제 될 수 있는 대표적 식물성 원료입니다. 구아 유래 갈락토만난은 물과 결합해 점성 네트워크를 만들 수 있고, 이 점성은 사료 혼합·소화효소 확산·장 내용물 이동에 영향을 줄 수 있습니다. 구아 meal을 섭취한 육계에서 효소와 발효 처리를 함께 평가한 연구는, 이 원료가 단순한 단백질 또는 섬유 공급원이 아니라 가공·소화 특성까지 고려해야 하는 기질임을 보여줍니다 [7].
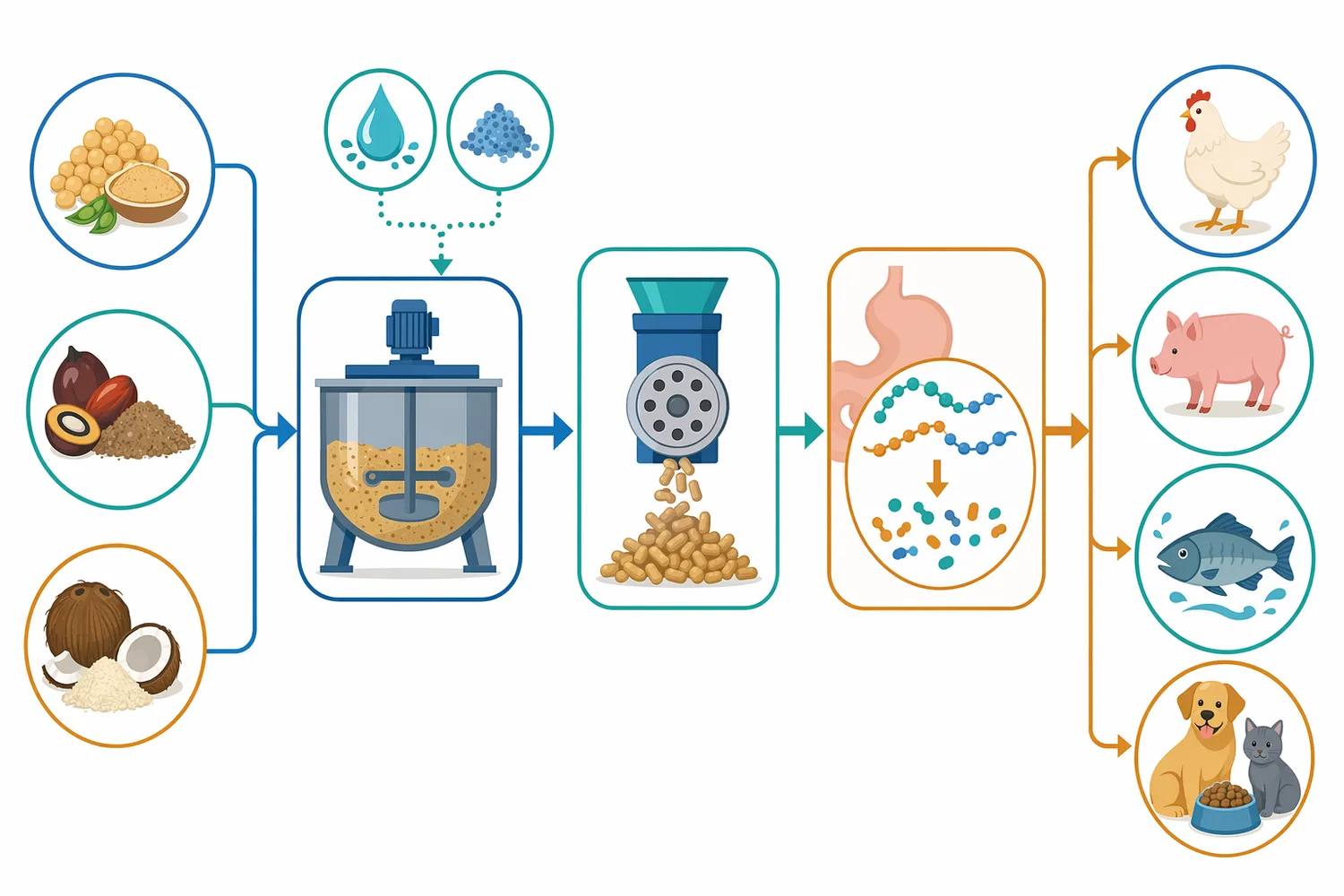
β-만나나아제는 갈락토만난의 만난 주사슬을 끊어 분자량을 낮추는 방식으로 접근합니다. 다만 갈락토스 측쇄가 많은 기질에서는 주사슬 접근성과 보조 효소 필요성이 달라질 수 있습니다. 따라서 구아 meal 같은 원료에서 기대 효과는 “만난성 점질 구조를 완화할 가능성”으로 표현하는 것이 적절하며, 모든 배합에서 동일한 성능 향상을 보장한다고 해석해서는 안 됩니다 [7].
Soy hulls는 섬유성 성분이 많아 사료 원료로 사용할 때 에너지 희석, 장내 체류, 영양소 이용성 변화가 함께 고려됩니다. 육계에서 soy hulls 수준과 β-mannanase 보충을 함께 평가한 연구는, 고섬유 원료를 단순히 제외하기보다 효소 보조를 통해 활용성을 조정하려는 접근이 실제로 검토되고 있음을 보여줍니다 [6].
그러나 soy hulls의 제한 요인은 만난만이 아닐 수 있습니다. 셀룰로오스, 펙틴, 자일란성 헤미셀룰로오스 등 다른 세포벽 성분도 함께 존재할 수 있으므로, β-만나나아제의 효과는 만난 분획이 전체 제한성에서 차지하는 비중에 따라 달라집니다. 식물 세포벽 분해 메커니즘 연구는 셀룰로오스 초분자 구조와 헤미셀룰로오스 매트릭스가 효소 가수분해를 함께 좌우한다는 점을 강조합니다 [9].
팜커널박이나 팜커널케이크 같은 부산물은 만난성 헤미셀룰로오스가 많은 원료로 자주 논의됩니다. 이러한 원료는 비용 측면에서 매력적이지만, 단위동물이나 일부 어류 사료에서는 높은 섬유성과 만난계 성분 때문에 사용 범위가 제한될 수 있습니다. 만난이 풍부한 바이오매스는 β-만나나아제 처리 후 만노올리고당 또는 발효성 당 전구체 생산으로 연결될 수 있다는 점에서 사료와 바이오리파이너리의 경계 영역에 위치합니다 [11].
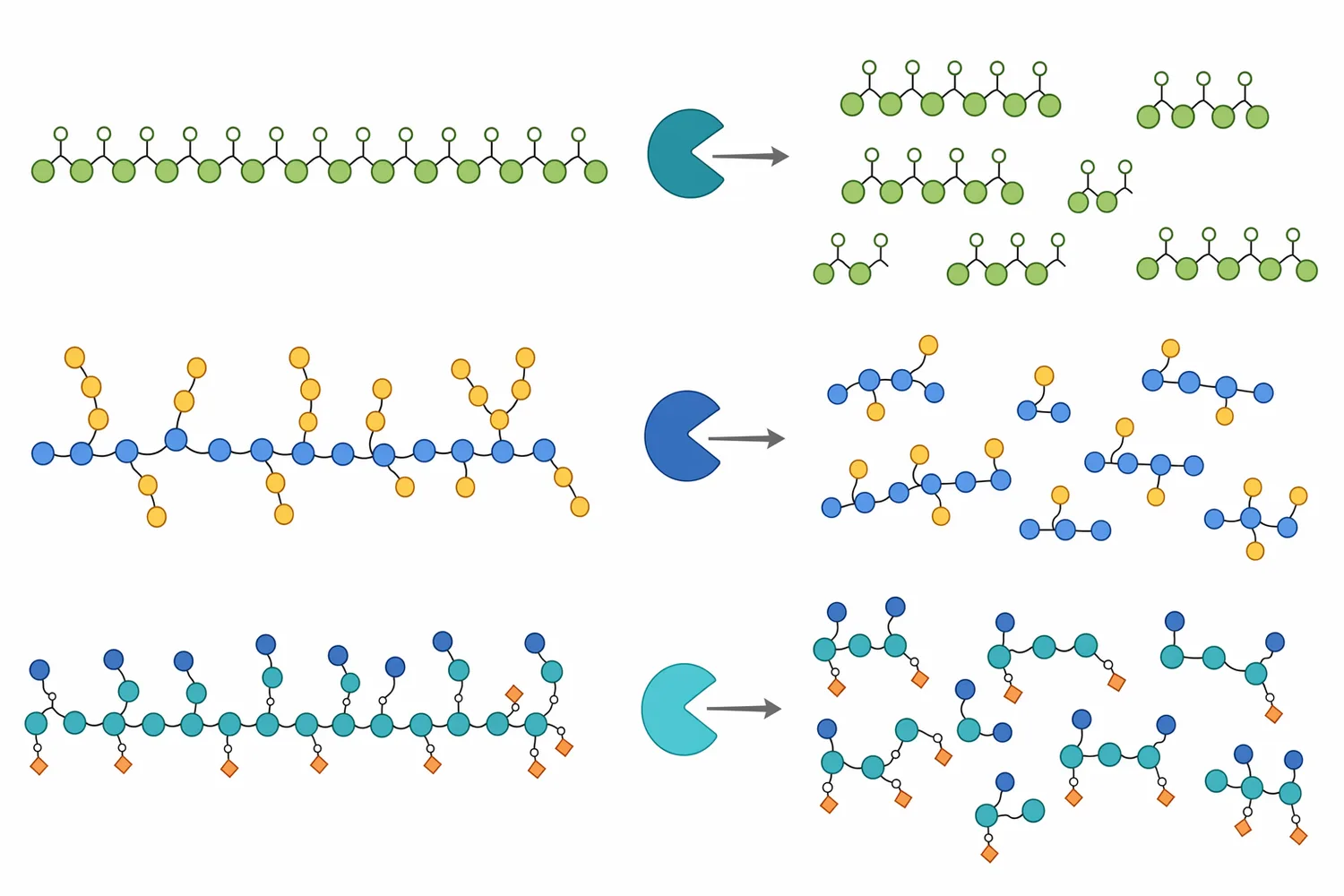
이 경우 β-만나나아제의 가치는 두 가지입니다. 첫째, 사료 원료로 사용할 때 만난성 항영양 요인을 줄이는 접근이 될 수 있습니다. 둘째, 원료 자체를 효소 처리해 MOS 또는 발효 가능한 당질 분획으로 전환하는 공정 소재가 될 수 있습니다. 다만 팜계 부산물은 리그닌과 다른 NSP도 포함하므로, 만난 분해만으로 전체 원료의 물성이 완전히 바뀐다고 기대하기는 어렵습니다 [5].
리그노셀룰로오스 바이오매스 전환은 셀룰로오스, 헤미셀룰로오스, 리그닌의 상호작용을 단계적으로 풀어내는 과정입니다. 효소가 어디에 결합하고, 어느 성분을 먼저 절단하며, 세포벽이 시간에 따라 어떻게 변형되는지가 전체 당화 효율을 좌우합니다. 싱크로트론 time-lapse imaging과 microfluidic infrared microspectroscopy를 활용한 연구는 효소 위치와 세포벽 화학 변화를 동시에 추적하려 했으며, 이는 바이오매스 가수분해가 단순한 “효소 투입” 이상의 구조적 과정임을 보여줍니다 [5].
β-만나나아제는 이 복합 공정에서 만난 분획을 담당합니다. 자일란이 많은 원료에서는 자일라나아제 계열, 셀룰로오스가 주요 탄소원인 원료에서는 셀룰라아제 계열, 펙틴이 큰 비중을 차지하는 원료에서는 펙티나아제 계열의 기여가 커질 수 있습니다. 미생물 xylanolytic enzyme system 연구가 여러 효소의 특성과 응용을 다루는 것처럼, 실제 식물 세포벽 처리에서는 표적 다당류별 효소 체계를 구분해 설계하는 관점이 중요합니다 [12].
또한 세포벽 성분 간 비공유 상호작용도 효소 반응을 좌우합니다. 최근 식물 세포벽 연구는 arabinogalactan protein O-glycans와 pectin 사이의 비공유 상호작용처럼, 다당류와 당단백질 네트워크가 벽 특성에 영향을 줄 수 있음을 보여줍니다. 이런 복합 상호작용은 만난 분해 효소가 잘 작동하더라도 전체 세포벽 붕괴가 제한될 수 있는 이유를 설명합니다 [13].
만노올리고당 생산에서는 완전한 당화보다 산물 분포가 중요합니다. β-만나나아제가 만난 사슬을 무작위에 가깝게 내부 절단하면 다양한 중합도의 올리고당이 생길 수 있고, 기질 구조와 효소 특성에 따라 짧은 MOS, 더 긴 MOS, 일부 만노스의 비율이 달라질 수 있습니다. 이 산물은 발효 미생물의 탄소원, 사료 내 기능성 당질, 또는 추가 효소 처리의 중간체로 검토될 수 있습니다 [11].
이때 “mannose oligosaccharide processing”이라는 표현은 세포 내 당단백질 가공과 혼동하지 않는 것이 중요합니다. 예를 들어 소포체와 골지체에서 일어나는 N-linked oligosaccharide processing은 glucosidase, mannosidase, glycosyltransferase 등이 관여하는 생합성·품질관리 경로이며, 사료 원료의 만난 가수분해와는 목적도 위치도 다릅니다 [14]. 본 제품의 맥락은 식물성 헤미셀룰로오스에서 유래한 만난 사슬을 효소적으로 절단하는 외부 공정입니다.
MOS 생산에서 β-만나나아제 단독 사용은 “올리고당 생성”에는 적합할 수 있지만, “고농도 만노스 생산”에는 충분하지 않을 수 있습니다. 만노스 단당을 목표로 한다면 β-만노시다아제 같은 말단 절단 효소나 발효 미생물의 대사능이 중요해집니다. 반대로 올리고당 자체가 목적이라면 과도한 후속 분해는 오히려 원하는 산물 분포를 무너뜨릴 수 있습니다 [4].
β-만나나아제는 사료 효율 개선 가능성을 가진 효소이지만, “모든 식물성 사료에서 동일하게 성장률을 높인다”는 식의 해석은 과학적으로 안전하지 않습니다. 돼지, 육계, 어류 연구에서 β-mannanase는 성장 성능, 영양소 이용, 분변 상태, 점도, 아미노산 소화율 같은 지표와 함께 평가되지만, 결과는 원료 조성, 동물 종, 성장 단계, 배합 영양 수준, 가공 조건에 따라 달라질 수 있습니다 [1].
특히 저만난 원료 중심의 사료에서는 β-만나나아제의 표적 기질이 충분하지 않을 수 있습니다. 반대로 구아 meal, soy hulls, 대두박 고함량 배합, 팜계 부산물처럼 만난성 비전분다당류가 의미 있게 존재하는 배합에서는 효소 적용 논리가 더 명확해집니다. 이 차이를 무시하면 효소가 실제로 작동할 기질이 부족한 상황에서도 성능 개선을 기대하는 오류가 생깁니다 [6].
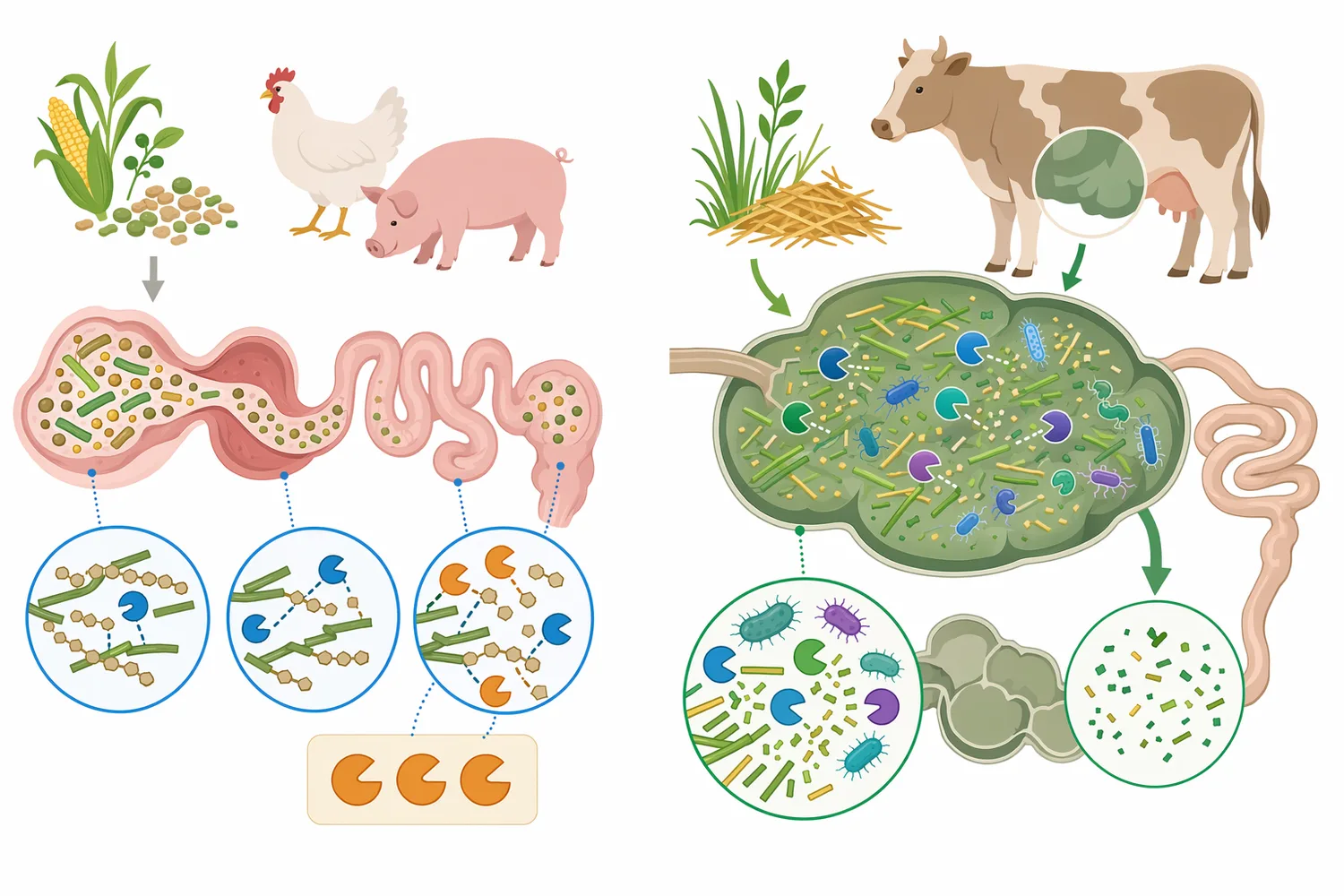
또한 외인성 효소는 영양 설계의 대체물이 아니라 보조 도구입니다. 에너지, 아미노산, 미네랄, 지방, 섬유 균형이 크게 어긋난 배합을 효소 하나로 보정하기는 어렵습니다. 최근 육계 연구에서도 영양소 밀도, 유화제, 소화율, 회장 아미노산 소화율처럼 여러 요소가 함께 평가되며, 사료 성능은 단일 첨가제보다 전체 배합 체계의 결과로 보는 것이 타당합니다 [15].
Enzymes.bio의 Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processing은 만난성 헤미셀룰로오스 처리와 사료 첨가 용도를 중심으로 이해하는 것이 적절합니다. 제품명에 포함된 feed addition, hemicellulose, mannose oligosaccharide processing은 각각 식물성 사료 원료의 β-만난 부담 완화, 세포벽 헤미셀룰로오스 표적 분해, MOS 또는 관련 당질 산물 생성 가능성을 가리키는 표현입니다 .
Enzymes.bio는 제조사 또는 분석 서비스 제공자가 아니라 공급업체입니다. 따라서 이 문서는 특정 제조 공정, 구체적 활성 단위, 분석법, 활성 정의를 설명하기 위한 자료가 아니라, 제품 페이지에서 효소의 기능적 의미와 문헌상 응용 범위를 이해하도록 돕는 기술 안내문입니다. 제품은 온라인에서 1kg 단위로 직접 구매할 수 있으며, 주문 시 CoA와 SDS가 함께 제공됩니다 .
실제 적용에서는 “이 원료에 만난계 헤미셀룰로오스가 충분히 존재하는가”가 가장 중요한 출발점입니다. 만난, 갈락토만난, 글루코만난이 주요 제한 요인인 공정에서는 β-만나나아제의 역할이 분명하지만, 주된 제한 요인이 자일란, 셀룰로오스, 리그닌, 피틴산 또는 단백질 소화 저해물질인 경우에는 다른 효소나 공정 접근이 더 직접적일 수 있습니다 [9].
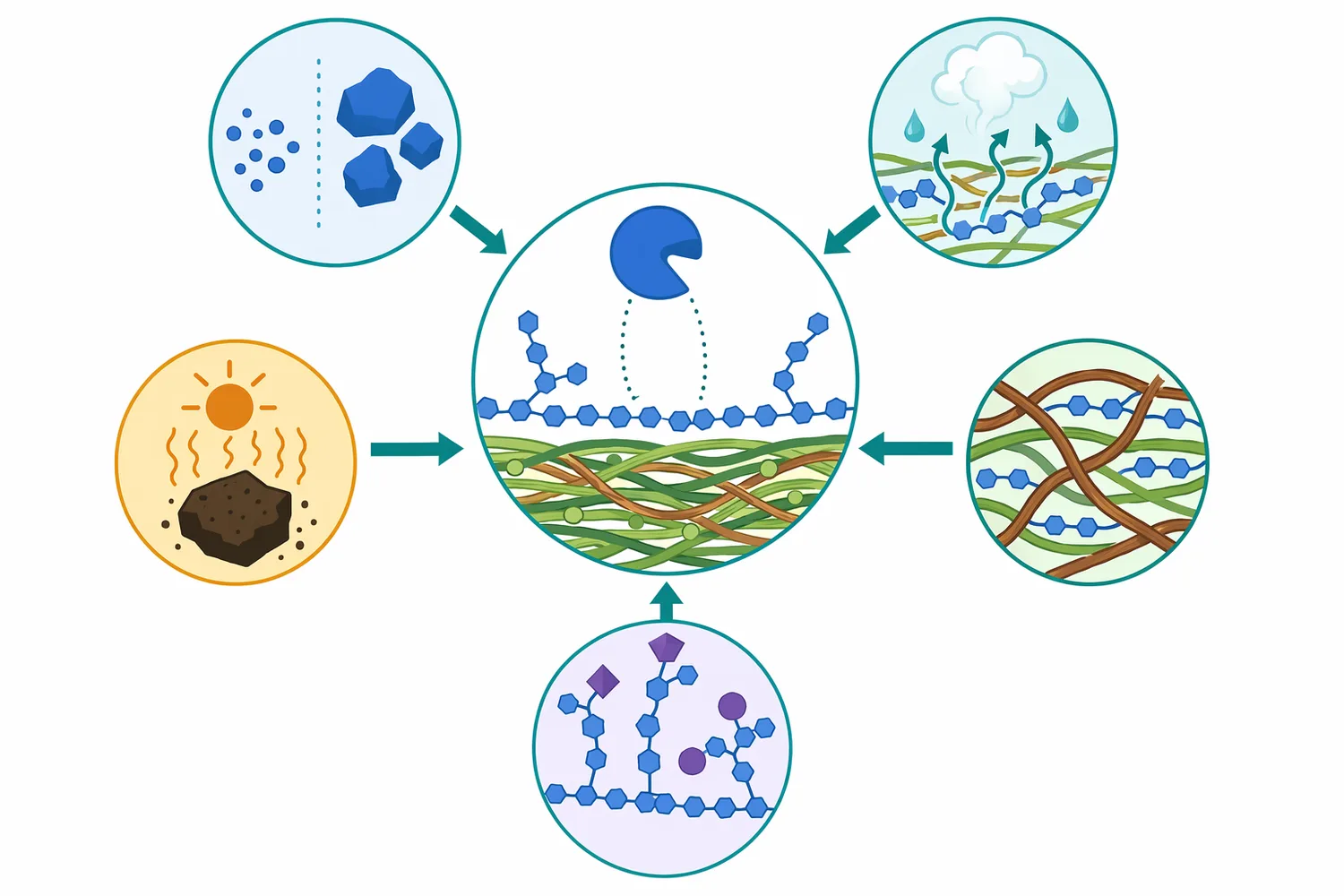
β-만나나아제는 식물성 만난계 헤미셀룰로오스의 β-1,4 결합을 절단해 긴 다당류를 더 짧은 만노올리고당과 관련 당질 산물로 전환하는 효소입니다. 이 반응은 사료에서는 β-만난성 비전분다당류의 점도·소화율·영양소 손실 문제를 완화하는 방향으로, 바이오매스 처리에서는 만난 풍부 원료의 MOS 또는 발효성 당 전구체 생산으로 연결될 수 있습니다 [2].
다만 β-만나나아제는 만난 표적 효소이지 리그노셀룰로오스 전체를 단독으로 완전 분해하는 만능 효소가 아닙니다. 복합 원료에서는 셀룰로오스, 자일란, 펙틴, 리그닌, 교차결합 구조가 함께 작용하므로, 효소의 효과는 기질 구성과 공정 목적에 따라 달라집니다 [4].
따라서 Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processing은 만난성 사료 원료, 구아·대두·soy hulls·팜계 부산물, 또는 만난 풍부 바이오매스의 표적 처리에 적합한 효소로 이해하는 것이 가장 정확합니다. Enzymes.bio에서는 해당 제품을 1kg 단위로 온라인 구매할 수 있으며, CoA와 SDS는 주문 시 함께 제공됩니다 .
1kg 단위로 판매되며 재고 보유, 즉시 출고됩니다. 온라인 스토어에서 바로 결제하시면 주문을 처리해 드립니다. 모든 주문에는 시험성적서(CoA)와 물질안전보건자료(SDS)가 포함됩니다.
Mannanase Feed Addition Hemicellulose Mannose Oligosaccharide Processing 구매하기 →최초 인용 순서로 번호를 매겼습니다. 모든 출처는 발행 시점에 접근 가능 여부를 확인한 오픈 액세스 자료이며, 본문의 인용 번호가 이곳으로 연결됩니다.