Neutral Protease Bacillus subtilis Protease is used to partially hydrolyze plant proteins into smaller peptides, improving how protein-rich materials disperse, solubilize, ferment, and function in ingredient systems. For soy, seed, gluten, and mixed plant-protein streams, the practical value is controlled protein modification: large, compact storage proteins are cut into shorter peptide fractions that are easier to handle in aqueous processing and can show improved functional properties.
Enzymes.bio supplies this neutral protease as an online 1 kg product for buyers who want a practical enzyme for plant protein hydrolysis; the order is paid for online, then processed and shipped, with a Certificate of Analysis and Safety Data Sheet included with the order. The product is positioned for plant protein hydrolysis applications rather than as a custom development service or laboratory testing program .
Neutral protease is a peptide-bond-cleaving enzyme used where the goal is not to destroy protein completely, but to modify it in a controlled way. In plant protein processing, that usually means reducing large storage proteins into peptide-rich hydrolysates with better water interaction, more accessible nitrogen fractions, and different functional behavior in emulsions, dispersions, fermentation media, or feed and food ingredient systems.
Plant proteins are often structurally difficult substrates. Soy proteins, wheat gluten, pea proteins, seed meals, and oilseed press cakes contain folded globulins, prolamins, glutelins, protein bodies, and protein–fiber associations that limit hydration and enzyme access. Neutral protease acts on accessible peptide bonds inside these structures, progressively loosening the protein network as peptide chains are shortened.
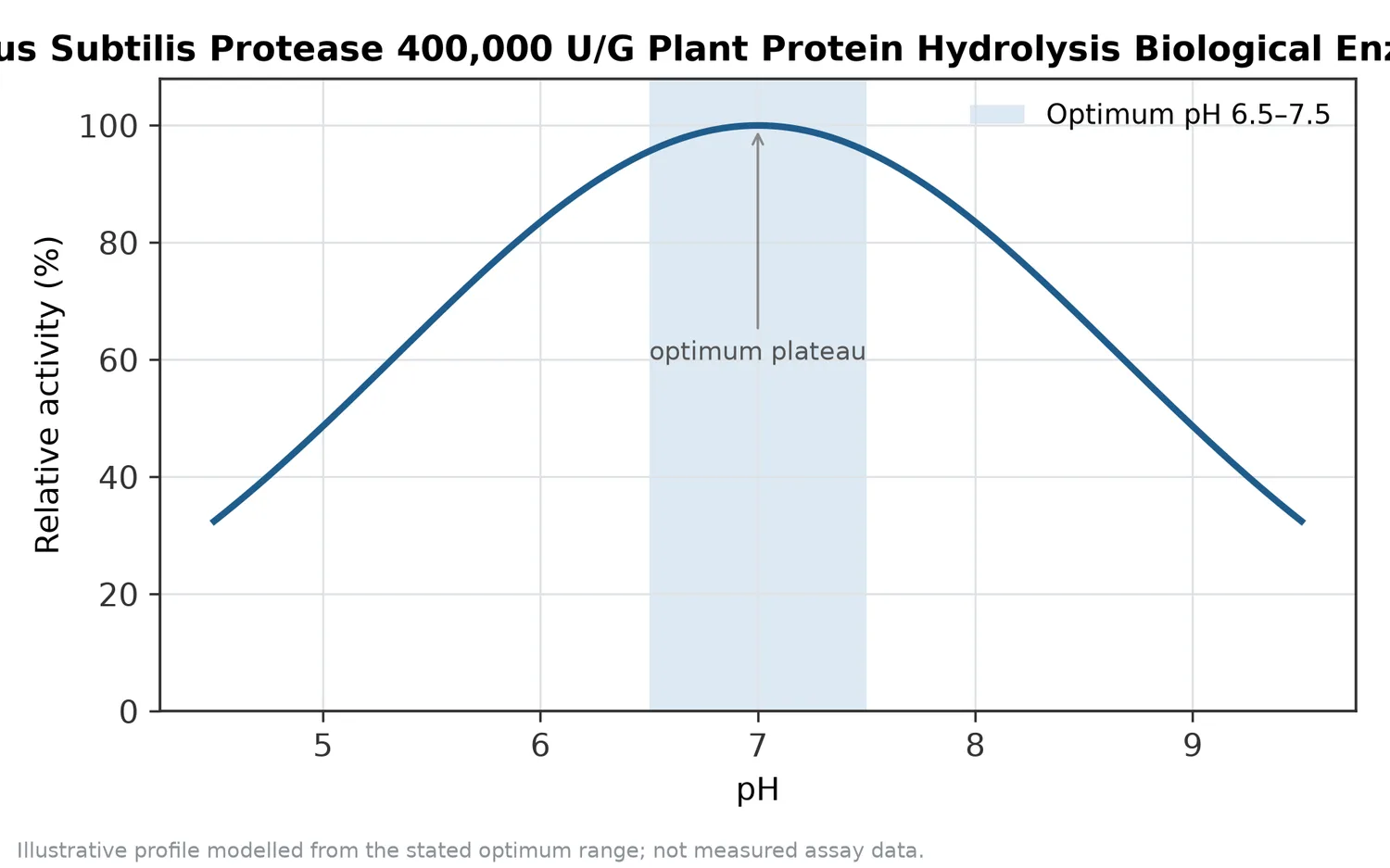
The term “neutral” is important because it distinguishes this enzyme class from acid proteases that work best in strongly acidic environments and alkaline proteases that are normally associated with higher-pH processes. Neutral proteases are especially useful where the product matrix, sensory profile, or downstream process benefits from milder conditions around neutral to mildly alkaline aqueous processing.
Bacillus subtilis is a well-studied microbial platform for extracellular enzyme production, including proteases used in food, feed, and industrial bioprocessing. Reviews of Bacillus species in food applications highlight their importance as enzyme producers and their long-standing relevance in fermented-food and ingredient systems, while also emphasizing the need to consider strain and application context [1].
A protein is a chain of amino acids folded into a three-dimensional structure. The peptide bonds are the backbone links between amino acids. Neutral protease catalyzes hydrolysis of those bonds by using water to split the chain, producing shorter peptides and, with sufficient hydrolysis, smaller amino-acid-containing fragments.
This molecular cutting has several practical consequences. Large proteins that were previously insoluble may become more dispersible because chain shortening exposes polar groups and reduces aggregation. Hydrophobic regions that were buried inside the folded protein may become exposed, changing emulsifying behavior. Charged amino acid side chains become more available to interact with water, salts, and other ingredients.
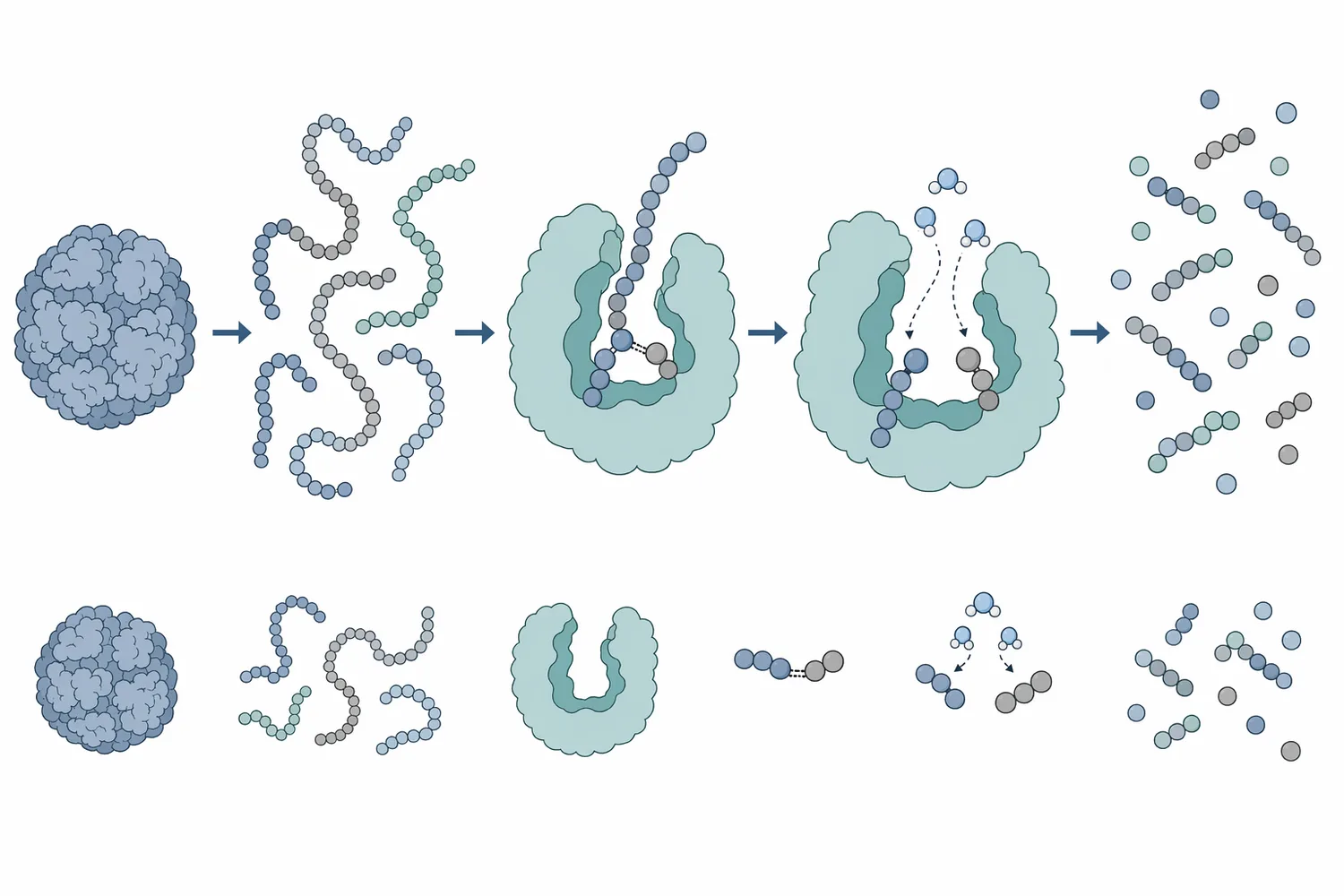
Hydrolysis also changes the balance between structure and mobility. A native soy globulin or gluten protein can behave like a large folded or network-forming molecule. After partial hydrolysis, the material contains many shorter peptides that move more freely in solution, diffuse faster, and interact differently with oil droplets, starch, fiber, minerals, and microbial cells.
Research on protein systems shows that protease treatment can shift the hydrophilicity–hydrophobicity balance of proteins, which matters because this balance controls whether protein fragments remain dispersed in water, adsorb at interfaces, aggregate, or contribute to foam and emulsion behavior [2]. In plant protein hydrolysis, the same principle explains why peptide size and surface chemistry strongly influence solubility, emulsification, and mouthfeel.
Many plant proteins have strong nutritional value but limited processing performance. Soybean proteins can contain compact storage fractions; wheat gluten is cohesive, elastic, and hydrophobic; seed proteins may be embedded in fiber-rich matrices; and plant meals may contain heat-denatured protein aggregates formed during oil extraction or drying.
Neutral protease helps by opening these structures gradually. Instead of relying only on heat, acid, alkali, or mechanical shear, enzymatic hydrolysis targets the protein backbone directly. That selectivity is valuable: the enzyme modifies protein while leaving starch, fiber, fat, and minerals largely outside its direct catalytic target.
In soybean protein isolate work, enzymatic hydrolysis by a neutral protease was investigated as a way to generate hydrolyzed soybean protein suitable as a precursor for thermal process flavours. This is a useful example because it connects hydrolysis to downstream ingredient functionality: peptides and amino-acid-rich fragments can participate differently in heat-driven flavor generation than intact soy proteins [3].
Neutral protease hydrolysis is also relevant to newer plant-protein ingredient development. Peptidomics research on soybean protein isolates has shown that neutral protease, alone or in combination with another protease, influences the release of antioxidant peptides from soy protein. The key point for processors is that the enzyme does not merely “dissolve protein”; it changes the peptide map, and that peptide profile determines functional and bioactivity-related outcomes [4].
Different protease classes can all hydrolyze protein, but they do not create the same processing experience or peptide profile. The table below gives a conceptual comparison for plant-protein applications without treating the categories as interchangeable specifications.
| Protease type | Typical processing character | Main practical effect on plant proteins | Where it can fit conceptually |
|---|---|---|---|
| Acid protease | Works best in acidic systems | Hydrolyzes proteins under low-pH conditions; may suit acidified slurries or certain fermentation-style matrices | Acid protein hydrolysates, acidic food systems, specialty processes |
| Neutral protease | Works around neutral to mildly alkaline aqueous conditions | Partially opens plant proteins while preserving a relatively mild process environment | Soy protein hydrolysates, seed protein hydrolysates, gluten modification, fermentation nutrients |
| Alkaline protease | Associated with higher-pH hydrolysis | Often gives strong protein breakdown and can be useful for more aggressive hydrolysis | High-protein waste valorization, detergent-like conditions, certain feed or industrial hydrolysates |
| Combined proteases | Uses complementary cleavage preferences | Produces broader peptide profiles because each enzyme cuts different peptide bonds | Bioactive peptide development, complex substrates, staged hydrolysis |
Protease selection changes peptide molecular weight distribution and resulting functional behavior. For example, research on oyster protein hydrolysates found that protease choice influenced molecular weight, antioxidant activity, and cellular protective effects, reinforcing the broader point that different proteases create different peptide populations even when acting on the same starting protein [5].
Soy is one of the most relevant plant substrates for neutral protease because it is widely used, protein-rich, and functionally complex. Soy protein isolate, soy flour, and soybean meal all contain storage proteins that can be difficult to hydrate or digest in their native or heat-treated states.
In soybean protein isolate hydrolysis, a neutral protease from Enterobacter sp. NRCM1 was used to produce enzymatically hydrolyzed soybean protein as a base precursor for thermal process flavours. Although the enzyme source in that study was not Bacillus subtilis, the work is directly relevant to neutral protease functionality because it demonstrates how neutral protease hydrolysis changes soy protein into a more reactive peptide-rich substrate for food ingredient applications [3].
Soy peptide research also shows why hydrolysis conditions and enzyme type matter. A peptidomics-based study of soybean protein isolates reported that neutral protease and Alcalase had synergistic effects on the release of antioxidant peptides. The study’s significance is not simply that hydrolysis occurred, but that the released peptide sequences and functional indicators depended on the enzyme system used [4].
For a buyer using Neutral Protease Bacillus subtilis Protease, the practical interpretation is straightforward: soy protein hydrolysis can be used to create peptide-rich materials with altered solubility, flavor-reactivity, and functional potential. The exact peptide profile will still depend on the soy raw material, pretreatment, hydrolysis endpoint, and downstream heating or drying.
Neutral protease research extends beyond soy. Milk thistle protein hydrolysis has been studied to compare the influence of protease type on structure and biological activity, showing that enzyme choice can alter structural features and activity indicators in seed-derived protein hydrolysates [6]. This matters because many plant proteins now used in foods and feeds come from oilseed and specialty seed streams rather than only soy.
Defatted winged bean-seed proteins provide another useful plant-protein example. Work on antinutritional factors and protease inhibitors in winged bean proteins described a denaturation, unfolding, and hydrolysis mechanism under thermal and hydrothermal treatments. Although that study focuses on treatment strategies rather than this specific commercial enzyme, it highlights an important processing reality: plant proteins often need unfolding or pretreatment before hydrolysis becomes efficient [7].
This is especially relevant for high-fiber or heat-processed raw materials. If proteins are trapped in dense particles or aggregated by prior heat exposure, enzyme access can be limited. Hydration, particle dispersion, and gentle unfolding can make peptide bonds more available to neutral protease, allowing the enzyme to work on the protein backbone rather than only on exposed surface regions.
Protein hydrolysates are often valued not only for solubility, but also for the biological or functional properties of their peptide fractions. Neutral protease hydrolysis of defatted Antheraea pernyi pupa protein has been reported to yield peptides with antioxidant activity, showing that neutral protease treatment can generate measurable activity in protein hydrolysates from dense protein substrates [8].
A separate study on silkworm pupa protein treated with a novel neutral protease also examined protein hydrolysates, again supporting the broader principle that neutral protease can convert intact protein into peptide fractions with changed functional or bioactivity-related properties [9]. These are not plant substrates, but they provide useful enzyme-mechanism evidence because the same catalytic principle—peptide bond hydrolysis—applies across protein-rich raw materials.
For plant protein applications, antioxidant peptide findings should be handled carefully. Laboratory antioxidant activity does not automatically become a finished-product health claim. It is best understood as product-development evidence: neutral protease can help generate peptide fractions worth evaluating for functional ingredient systems, but final claims depend on the actual finished product and applicable regulations.
The most defensible commercial benefit of neutral protease in plant proteins is improved functionality. This means the hydrolysate behaves differently from the original protein in water, oil–water systems, thermal processes, and fermentation.
Solubility can improve because smaller peptides expose more polar and charged groups per unit mass. Intact plant proteins often aggregate through hydrophobic patches, disulfide-linked structures, or heat-induced associations. Once neutral protease cuts those chains, the fragments may have less ability to form large insoluble aggregates and more ability to remain dispersed.
Emulsifying behavior can also change. Peptides with both hydrophobic and hydrophilic regions can migrate to oil–water interfaces and help stabilize droplets. Too little hydrolysis may leave proteins too large and insoluble; too much hydrolysis may produce fragments too small to build strong interfacial films. This is why controlled hydrolysis, rather than maximum hydrolysis, is usually the more useful processing goal.
In flavor systems, hydrolysis increases the pool of peptides and amino-acid-containing fragments available for thermal reactions. The soybean protein isolate study on hydrolysates for thermal process flavours is relevant here because it treats enzymatically hydrolyzed soy protein as a precursor material for downstream heat-generated flavor chemistry [3].
Soybean proteins such as glycinin and β-conglycinin are important in feed and food discussions because they can influence digestibility and antigenic response in sensitive applications. Protease treatment is often considered because breaking these proteins into smaller fragments can reduce recognition of intact antigenic structures.
However, it is important not to overstate what a neutral protease can guarantee. Hydrolysis can reduce intact protein fractions and may reduce antigenicity indicators under controlled conditions, but it does not automatically make a soy-containing material allergen-free. Allergenicity depends on remaining epitopes, processing history, target population, and regulatory context.
Research on cow milk protein antigenicity shows the same principle in a different protein system: different proteases affect hydrolysis degree and antigenicity differently, meaning enzyme choice and process control strongly influence the final antigenic profile [10]. The lesson for soy and other plant proteins is that protease hydrolysis is a useful modification tool, not a universal allergen-elimination claim.
For feed-oriented plant proteins, hydrolysis may also support digestibility by reducing large storage proteins into smaller peptides. Still, digestibility depends on the animal species, diet matrix, heat history, fiber content, and non-protein antinutritional factors. Neutral protease targets proteins; it does not directly remove phytate, fiber, tannins, or oligosaccharides.
Soy protein isolate, concentrate, flour, and meal are natural candidates for neutral protease treatment. Hydrolysis can support better dispersion, altered viscosity, improved peptide availability, and preparation of protein hydrolysates for food, feed, fermentation, and flavor applications.
Soybean peptide research demonstrates that neutral protease can contribute to the release of antioxidant peptide fractions from soy protein isolate, especially when combined with complementary protease activity [4]. In practical use, this supports neutral protease as a tool for creating differentiated soy hydrolysates rather than simply as a generic protein-degrading additive.
Wheat gluten is cohesive and hydrophobic, making it difficult to use in many aqueous ingredient systems. Neutral protease can partially cut gluten proteins into smaller fragments, reducing excessive network strength and improving dispersion potential.
Although the verified source list here is stronger for soy and seed proteins than for wheat gluten specifically, the mechanism is directly applicable: gluten functionality is driven by protein chain length, aggregation, and hydrophobic interactions. Protease hydrolysis changes those variables by cutting peptide bonds and shifting the balance between large elastic proteins and smaller soluble peptides.
Oilseed meals contain valuable protein but are often physically complex because protein is associated with fiber, residual oil, minerals, and heat-treated cell-wall material. Neutral protease can help unlock protein value when the process allows sufficient hydration and enzyme contact.
The winged bean-seed protein work is useful in this context because it emphasizes how denaturation and unfolding can be coupled with hydrolysis to reduce problematic protein factors and improve access to the substrate [7]. This aligns with real plant-meal processing, where protein accessibility is often as important as enzyme activity.
Protease-treated plant proteins can serve as nitrogen sources in fermentation. Shorter peptides and amino-acid-containing fragments are generally more accessible to microbes than intact storage proteins, particularly when the starting protein is poorly soluble.
Bacillus subtilis itself is widely studied for high-value bioproduction, including enzyme and metabolite applications, which reinforces the broader relevance of Bacillus-linked enzyme systems in fermentation-oriented processing [11]. For a buyer, the key benefit is practical: neutral protease can help convert plant protein into a more fermentable nitrogen fraction.
In feed applications, plant protein hydrolysis may help improve the nutritional value of soy meal, seed meals, and other protein-rich streams by reducing intact storage proteins and increasing peptide availability. This is particularly relevant where young animals or sensitive species respond poorly to poorly digested protein fractions.
The same caution applies: neutral protease modifies protein, not the entire antinutritional profile. If the raw material contains non-protein constraints, separate processing steps may still be needed. Neutral protease is best viewed as one protein-focused tool within a broader feed-ingredient process.
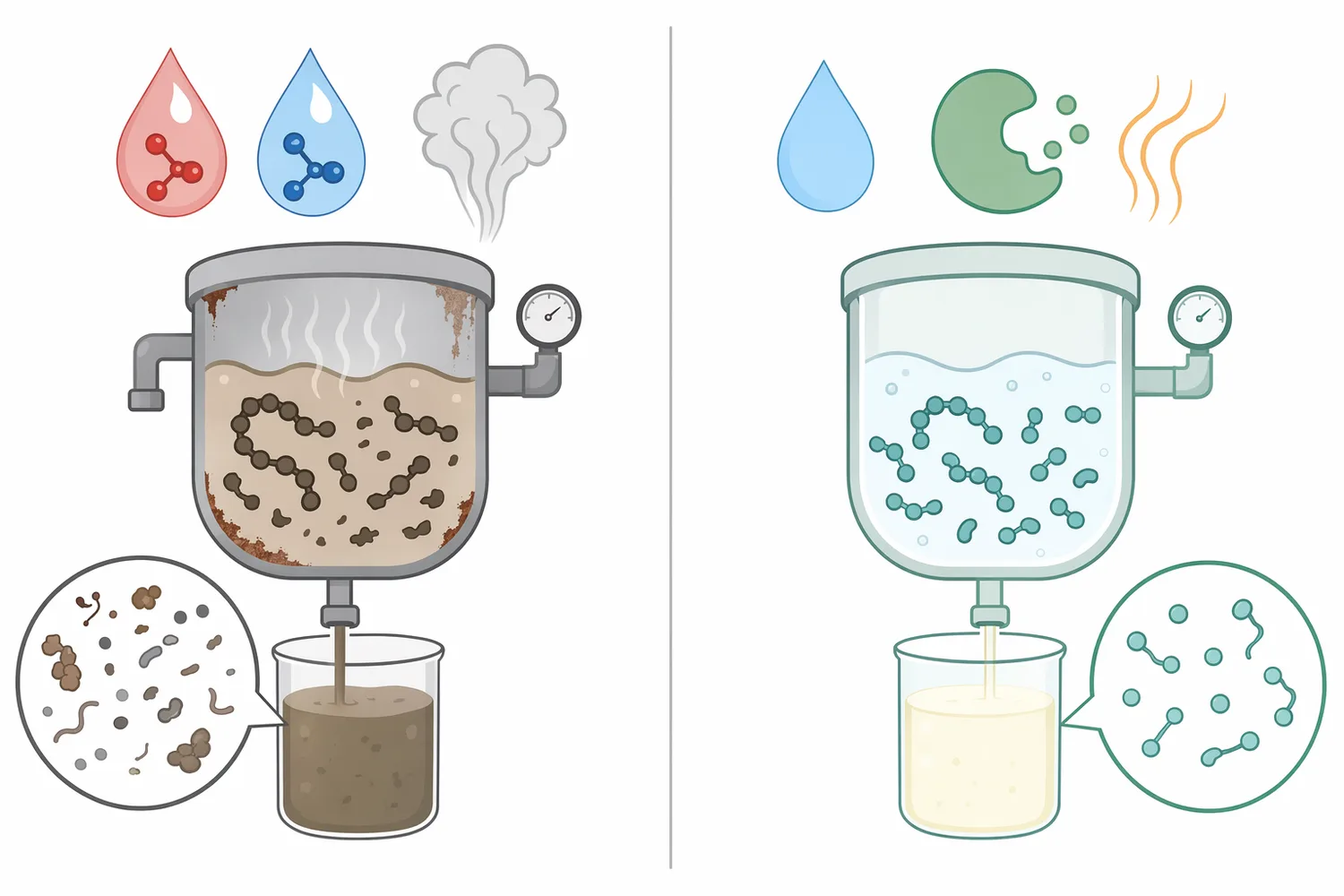
Neutral protease works in water because hydrolysis requires water as a reactant. In a plant-protein slurry, the enzyme must first reach the protein surface, bind accessible peptide regions, and catalyze chain cleavage. Good dispersion matters because dry clumps, oil-coated particles, or dense fiber-protein aggregates reduce contact between enzyme and substrate.
As hydrolysis progresses, the slurry may become easier to mix if large protein networks are broken down. In some systems, viscosity can fall as protein chains shorten. In others, exposed hydrophobic regions may temporarily increase aggregation until further hydrolysis produces more soluble fragments. This is why plant protein hydrolysis is often nonlinear: the first stage opens the protein, and later stages reshape the peptide population.
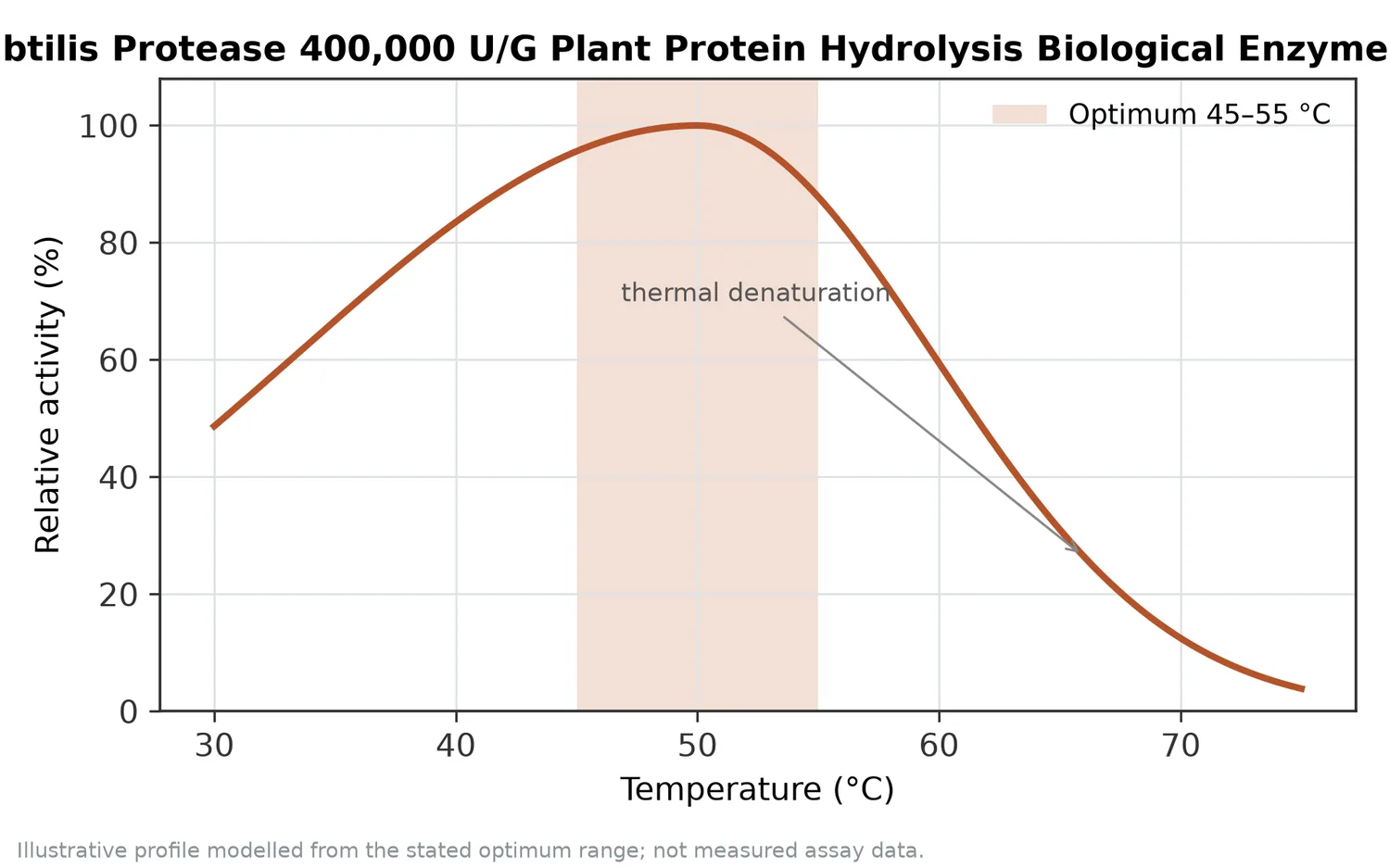
Temperature and pH influence enzyme conformation and substrate structure. Neutral protease is used in mild aqueous processes because extreme acid or alkali is not the point of the application. The practical operating window should be confirmed within the buyer’s own process, but the conceptual advantage remains the same: hydrolysis can be performed without relying on harsh chemical cleavage.
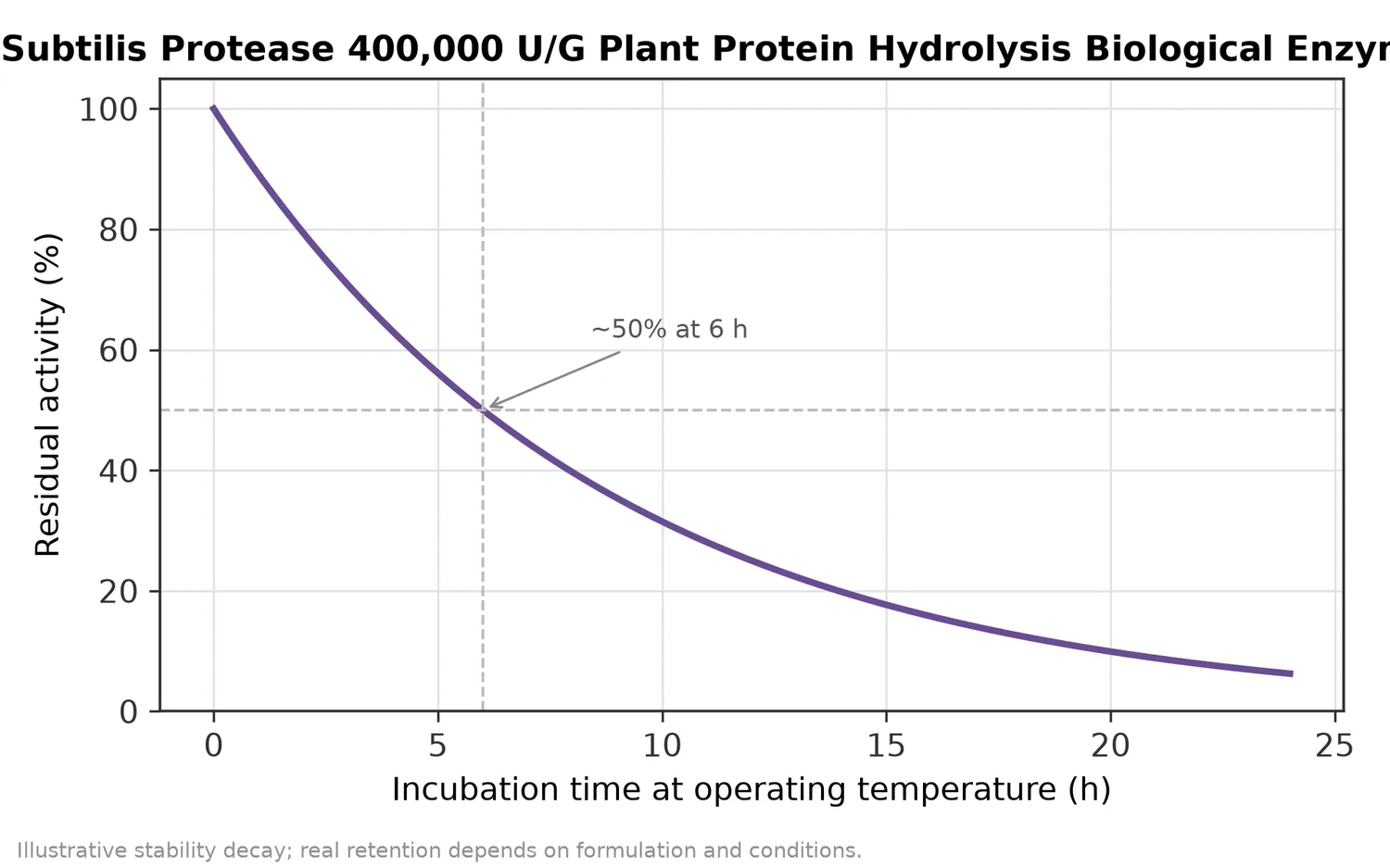
Enzyme inactivation is normally part of a controlled hydrolysis process. Once the desired endpoint is reached, heat or another suitable processing step stops further protease action so that the hydrolysate does not continue changing during downstream handling. The appropriate inactivation approach depends on the finished matrix and process design.
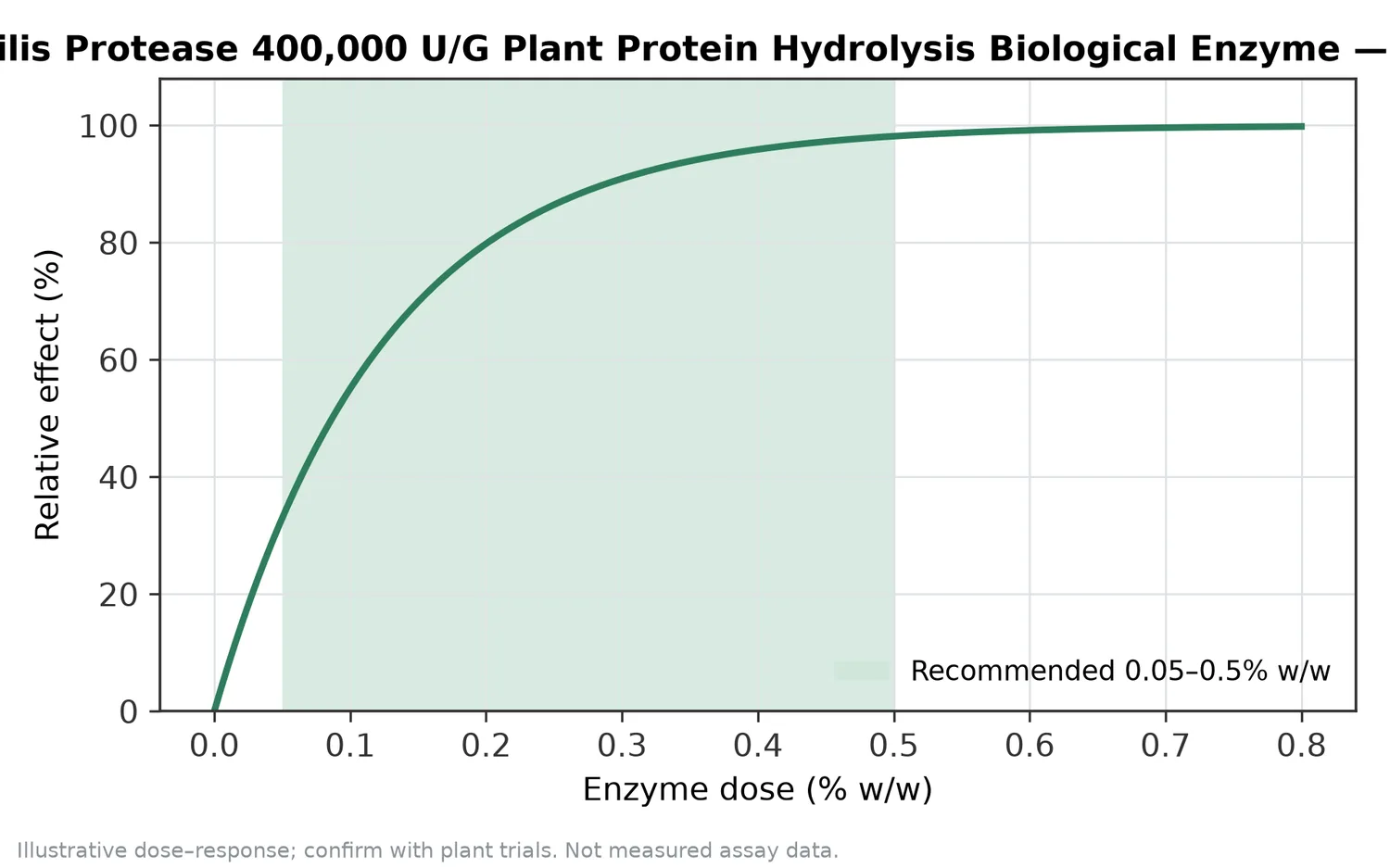
Partial hydrolysis is often more useful than complete hydrolysis. A lightly hydrolyzed plant protein may show better solubility while still retaining body, emulsifying structure, or mild flavor. A more extensively hydrolyzed protein may be more soluble but can become bitter, thinner, or more reactive during heating.
Bitterness is a common issue in protein hydrolysates because some hydrophobic peptides taste bitter. Neutral protease can expose or release such peptides depending on the substrate and hydrolysis extent. This does not make hydrolysis unsuitable; it means the hydrolysis endpoint should match the intended use, whether that is a savory base, feed ingredient, fermentation nutrient, beverage component, or functional powder.
Peptide size also affects drying and reconstitution. Smaller peptides can dissolve faster, but they may also increase hygroscopicity or alter powder flow. Larger peptides may contribute more body but disperse less completely. These outcomes are material-specific, which is why the same neutral protease can produce different handling behavior in soy isolate, soybean meal, pea protein, and gluten.
Studies comparing protease types across protein substrates consistently show that enzyme choice affects molecular weight distribution and functional indicators. The oyster hydrolysate study is a clear example outside plant proteins, demonstrating that protease selection changed molecular weight and antioxidant outcomes in the resulting hydrolysates [5].
Neutral protease does not hydrolyze carbohydrates, fats, phytate, or most non-protein antinutritional factors. Its catalytic target is the peptide bond. If a plant meal’s limitation is primarily fiber, phytate, starch gelatinization, or lipid oxidation, protease may help the protein fraction but will not solve the entire material challenge.
It also does not guarantee allergen elimination. Even when intact proteins are reduced, peptide fragments may still contain epitopes relevant to sensitive individuals. Any allergen-related positioning must be supported by product-specific testing and regulatory review.
Neutral protease does not guarantee a particular flavor. Hydrolysis can improve savory depth or thermal flavor reactivity in some applications, but it can also release bitter peptides. The soybean protein hydrolysate work for thermal process flavours supports the use of hydrolysates as flavor precursors, but the final flavor depends on the reaction system, heating conditions, reducing sugars, amino compounds, salt, fat, and other formulation factors [3].
Finally, neutral protease does not produce the same hydrolysate from every plant protein. Protein sequence, folding, pretreatment, heat history, particle size, and matrix composition all influence which peptide bonds are accessible and which peptides accumulate.
Enzymes.bio supplies Neutral Protease Bacillus subtilis Protease for buyers who want to purchase the 1 kg product directly online and use it in their own plant protein hydrolysis work. The buyer pays online, the order is processed and shipped, and the accompanying Certificate of Analysis and Safety Data Sheet support internal receiving, documentation, and safe handling.
This product fit is straightforward: it is for teams that already have a plant-protein process or development workflow and need a neutral protease ingredient to evaluate or use in hydrolysis. Enzymes.bio is the supplier of the product, not a contract laboratory or custom manufacturing service.
The best-supported applications are soy and other plant protein hydrolysates, peptide-rich ingredient systems, fermentation nutrients, feed protein modification, and improved use of protein-rich side streams. The underlying science is consistent across these uses: neutral protease cuts peptide bonds, reduces protein size, changes solubility and interfacial behavior, and creates new peptide profiles.
Neutral Protease Bacillus subtilis Protease is a practical enzyme for converting dense plant proteins into more usable peptide-rich hydrolysates. Its main value is controlled protein modification: improving dispersion, solubility, peptide availability, and functional behavior without relying on harsh chemical hydrolysis.
The evidence base supports neutral protease hydrolysis for soy protein systems, seed proteins, peptide generation, and functional hydrolysate development, while broader Bacillus literature supports the relevance of Bacillus subtilis as an important enzyme-producing organism in food and industrial biotechnology [1]. For buyers using Enzymes.bio’s online 1 kg product, the realistic expectation is not a universal one-step solution, but a dependable protein-hydrolysis tool for plant-protein processing.
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Neutral Protease Bacillus Subtilis Protease 400,000 U/G Plant Protein Hydrolysis Biological Enzyme →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.