A nuclease is an enzyme that cleaves nucleic acids—DNA, RNA, or both—by breaking the phosphodiester bonds that connect nucleotide units. In practical use, nuclease function is to convert long DNA or RNA molecules into shorter fragments, which can reduce nucleic-acid burden, improve cleanup, and make downstream handling easier. Enzymes.bio supplies Nuclease directly online by the 1 kg unit; after online purchase, the order is processed and shipped with a Certificate of Analysis and Safety Data Sheet.
The simplest nuclease definition is: an enzyme that cuts nucleic acid chains. DNA and RNA are long polymers built from nucleotides, and each nucleotide is linked to the next through a phosphodiester bond in the sugar-phosphate backbone. What a nuclease does is chemically specific: it hydrolyzes one or more of those backbone bonds, so a long strand becomes shorter fragments rather than remaining as an intact, high-molecular-weight polymer.
That nuclease meaning covers a broad enzyme category, not one single enzyme with one single behavior. Some nucleases act mainly on DNA, some on RNA, and some on both; some cut inside the chain, while others digest from an exposed end. This is why “what is a nuclease?” has a short answer—an enzyme that degrades DNA/RNA—and a more technical answer involving substrate preference, cleavage position, structure, and biological role.
Nucleases are fundamental in biology because cells constantly need to process nucleic acids. DNA repair systems use nucleases to remove damaged sections; RNA-processing pathways use ribonucleases to shape or degrade RNA; immune and defense systems use nuclease activity to destroy foreign genetic material. Studies of DNA repair nucleases such as UvrABC and FAN1 show that nuclease activity can be highly organized around damaged or abnormal DNA structures rather than being random chemical degradation [1].
For applied users, the important point is practical: nuclease is used where DNA or RNA is unwanted, excessive, structurally disruptive, or analytically interfering. In a biological mixture, intact genomic DNA can be large, charged, and physically persistent; cutting it into shorter pieces changes how it behaves in solution and how easily it can be separated from the material of interest.
A nuclease acts at the sugar-phosphate backbone of DNA or RNA. The enzyme binds the nucleic-acid substrate, positions a phosphodiester bond in its catalytic site, and promotes hydrolysis of that bond. The result is cleavage of the strand into smaller pieces. Structural work on staphylococcal nuclease used a thymidine 3′,5′-bisphosphate–calcium ion complex at 1.5 Å resolution to propose how the enzyme positions a nucleotide-like substrate and metal ion chemistry during catalysis [2].
This mechanism is concrete, not just a generic “breakdown” claim. DNA and RNA are stable because their phosphodiester bonds form a continuous backbone. A nuclease changes the molecule by interrupting that backbone. Once a few internal cuts are made, a long DNA molecule that previously behaved like an extended polymer becomes a collection of shorter fragments. Those fragments have lower chain length, fewer opportunities for entanglement, and different separation behavior in filtration, clarification, chromatography, or analytical preparation.
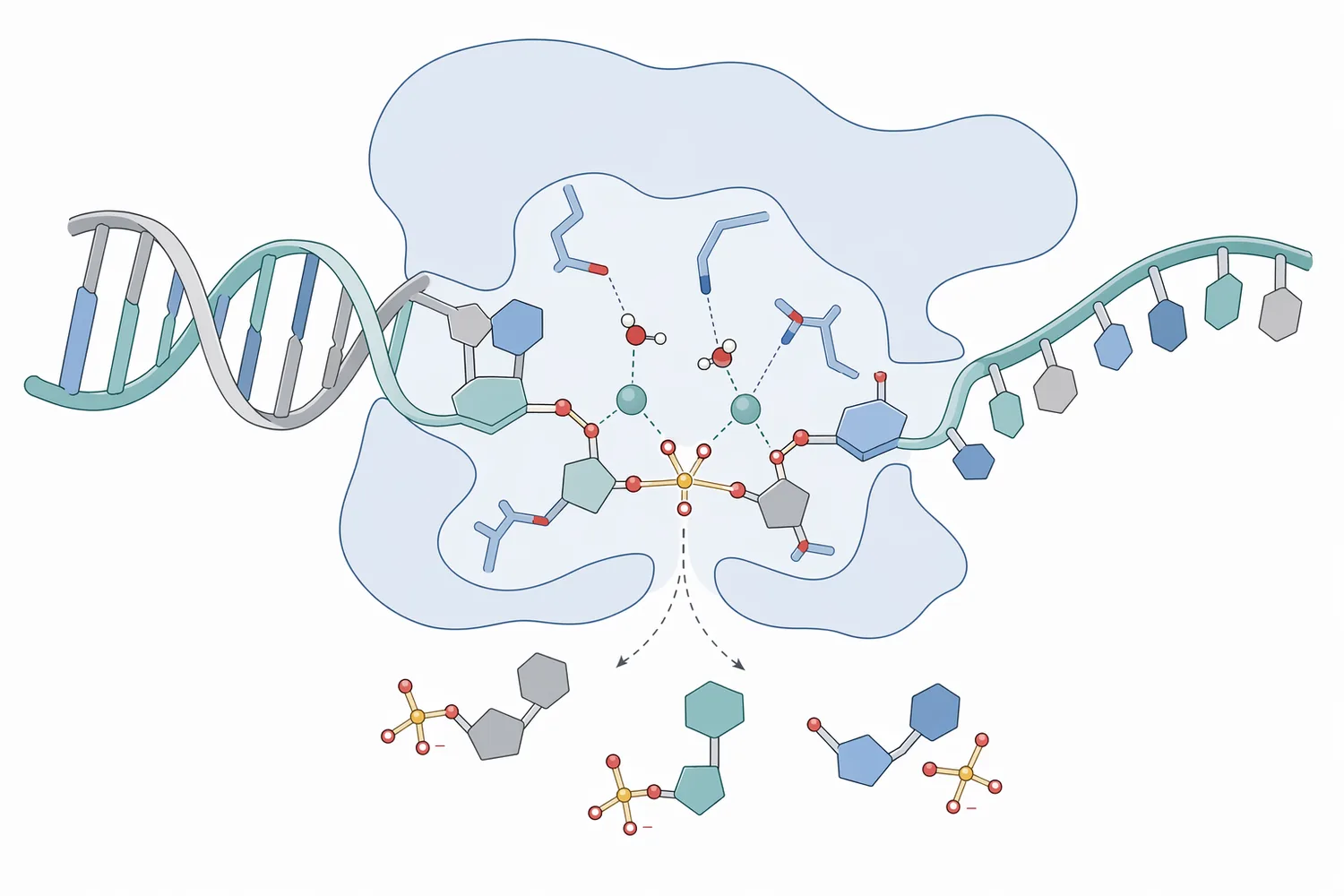
Many nucleases use metal ions to assist catalysis. The metal ion can help organize the phosphate group, activate water for attack, and stabilize charged transition states during bond cleavage. High-resolution structural analysis of Serratia marcescens nuclease at 1.7 Å provided a detailed basis for understanding how this broad nuclease family recognizes and cleaves nucleic-acid substrates [3].
Different nucleases also differ in where they cut. Endonucleases cut within a nucleic-acid strand. Exonucleases remove nucleotides or short units from an end. This difference matters because internal cutting can rapidly reduce the length of long DNA, while end-digestion produces a more stepwise trimming pattern. BAL 31 nuclease studies, for example, examined exonuclease action and kinetics on small substrates and single-stranded DNA, showing that nuclease behavior depends on substrate form and cleavage mode [4].
“Nuclease” includes many named enzymes and enzyme families. The table below gives a conceptual comparison without turning the category into a specification checklist.
| Nuclease type or example | Main concept | What changes in the substrate | Typical technical context |
|---|---|---|---|
| DNase | Acts on DNA | DNA backbone is cut into shorter DNA fragments | DNA removal, viscosity reduction, DNA cleanup |
| RNase | Acts on RNA | RNA strand is degraded or processed | RNA removal, RNA biology, sample preparation |
| Endonuclease | Cuts inside a strand | Long chains are internally fragmented | Fast reduction of intact nucleic-acid length |
| Exonuclease | Digests from an end | Chain ends are progressively shortened | Controlled trimming, end-processing, specialized digestion |
| S1 nuclease / nuclease S1 | Known for activity on single-stranded nucleic-acid regions | Single-stranded DNA or RNA regions can be preferentially cleaved | Molecular biology and nucleic-acid structure work |
| Micrococcal nuclease | Broad nuclease historically used to degrade DNA in biological extracts | DNA is degraded into smaller fragments under suitable conditions | Chromatin, extract preparation, nuclease-treated systems |
| Serratia-type nuclease / Benzonase nuclease category | Broad-spectrum nuclease concept associated with nonspecific DNA/RNA degradation | DNA and RNA can be fragmented rather than remaining as long polymers | Bioprocess cleanup and nucleic-acid burden reduction |
| Programmable nuclease: zinc finger nuclease, transcription activator-like effector nuclease / TALE nuclease, CRISPR-associated nuclease | Engineered or guide-directed DNA cutting | A defined genomic site is cut, rather than bulk nucleic acid being degraded | Genome engineering, gene disruption, targeted editing |
The distinction between general nucleic-acid degradation and sequence-directed cutting is especially important. A zinc finger nuclease or transcription activator-like effector nuclease—often written as TALE nuclease or TALEN—is designed for targeted DNA cleavage. A general nuclease used for DNA/RNA breakdown is normally intended to reduce nucleic-acid burden, not to edit a specific genome sequence.
Micrococcal nuclease is a useful example of how long-established nuclease tools support biological workflows. Early work on micrococcal nuclease studied degradation of thymus deoxyribonucleic acid, demonstrating its ability to act directly on DNA substrates [5]. Later biological studies also used micrococcal nuclease-treated cell-free extracts, reflecting how nuclease treatment can change the nucleic-acid content of a complex biological system [6].
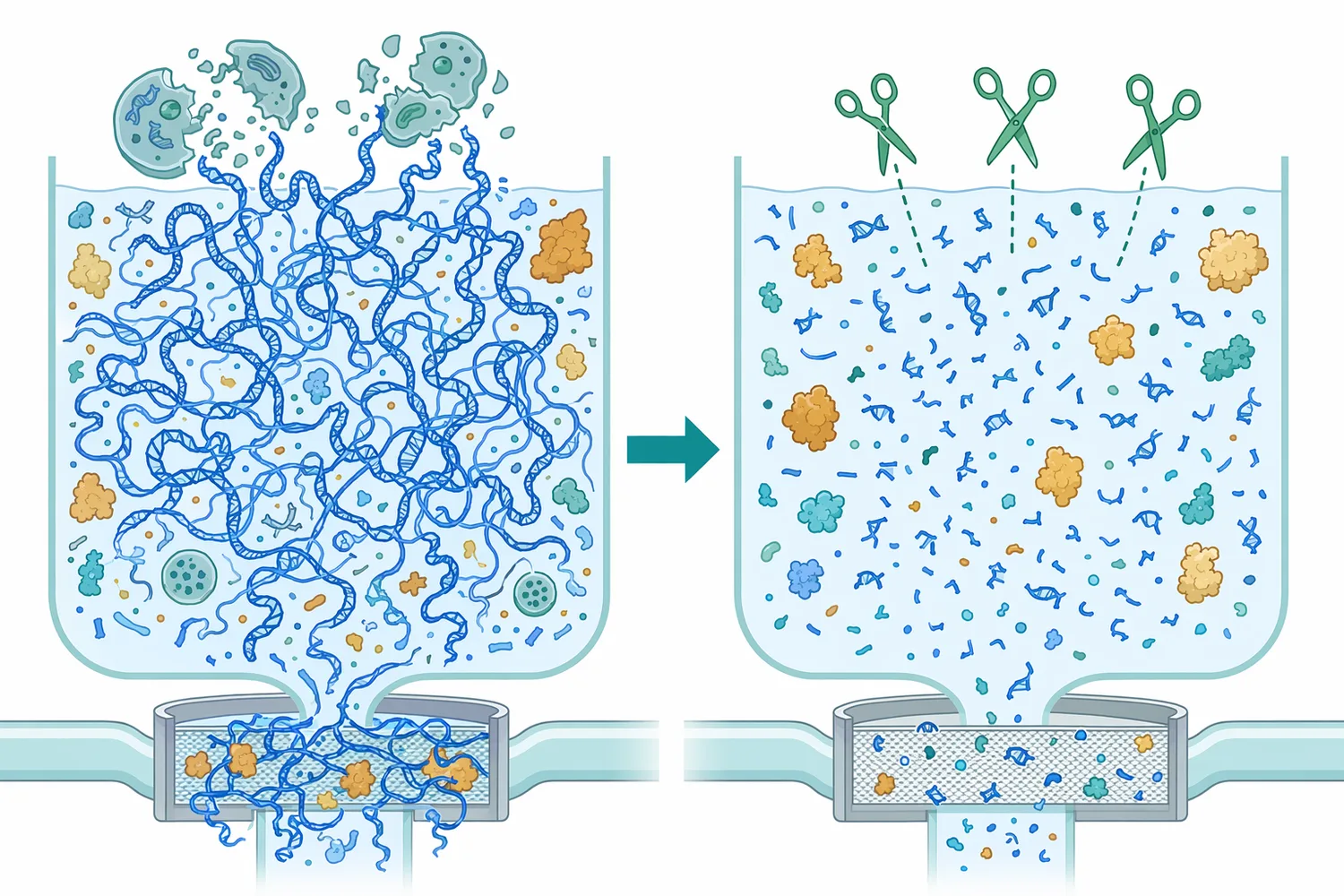
DNA and RNA are not neutral, inert background materials. They are large, highly charged polymers. When cells are disrupted, genomic DNA and RNA can enter the surrounding liquid and change its physical and chemical behavior. Long DNA is especially problematic because it can contribute to viscosity, bind proteins and surfaces electrostatically, and persist through process steps unless fragmented.
The mechanism of improvement is straightforward: nuclease does not make DNA disappear instantly; it changes DNA size. By cutting the backbone, it converts large molecules into shorter fragments. Shorter fragments are typically easier for downstream operations to handle because they are less polymeric, less entangling, and more separable than intact chromosomal DNA.
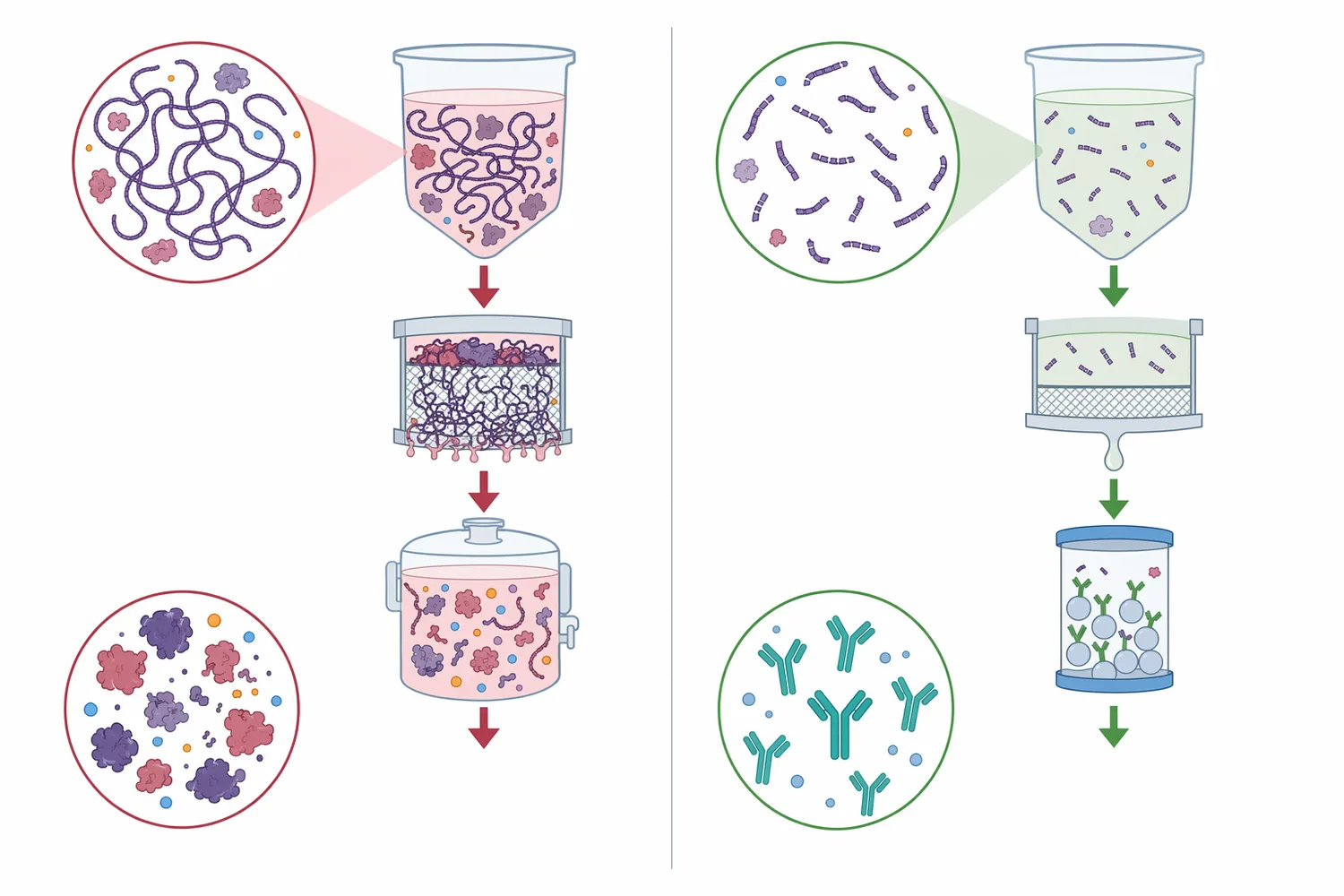
This is directly relevant in biological production and cleanup workflows. In cell-derived materials, nucleic acids are released together with proteins, membranes, chromatin components, and other cell contents. A nuclease treatment step can reduce the nucleic-acid contribution before or during later separation steps, helping the process deal with smaller fragments rather than long, intact DNA.
Nuclease also matters in analytical contexts. Residual DNA or RNA can interfere with assays, sample interpretation, or molecular biology work. That is why technical users often search for terms such as “nuclease-free water,” “water nuclease free,” or branded phrases like “nuclease free water Ambion.” Those searches are about preventing unwanted nuclease contamination during nucleic-acid work—the opposite problem from intentionally adding nuclease to degrade DNA or RNA.
In bioprocessing, nuclease is commonly discussed where host-cell nucleic acids create a cleanup burden. Cell disruption releases DNA into a mixture that may also contain the desired protein, viral vector, enzyme, or other biological product. Because DNA is long and negatively charged, it can associate with proteins, chromatin material, and process surfaces; fragmentation helps downstream steps deal with a smaller and less physically disruptive contaminant.
The key value is not that nuclease performs a complete purification by itself. Its value is that it changes the contaminant before separation. A long DNA molecule may behave like a sticky, high-molecular-weight polymer; after nuclease action, the same genetic material exists as shorter pieces. That molecular change can support clarification, filtration, and other downstream operations by reducing the effects of intact nucleic acid.
Salt and matrix chemistry can influence this process because nuclease-substrate binding often depends on charge interactions between the enzyme and the nucleic-acid backbone. Industry discussion of salt-active nucleases in biopharmaceutical manufacturing highlights that some process environments contain salt levels that can challenge conventional nuclease performance, while specialized nuclease approaches are discussed for compatibility with such conditions [7].
This chemistry matters because DNA is negatively charged. When ionic strength increases, dissolved ions can shield electrostatic attractions between enzyme surfaces and DNA, making it harder for some nucleases to bind and cut efficiently. Conversely, in a compatible environment, the nuclease can access the DNA backbone, make repeated cuts, and reduce polymer length.
The nuclease enzyme category is supported by decades of structural and biochemical research. Staphylococcal nuclease is one of the classic systems for understanding enzyme-substrate recognition and metal-assisted phosphodiester cleavage. The 1.5 Å structure of a staphylococcal nuclease complex with thymidine 3′,5′-bisphosphate and calcium ion provided atom-level evidence for how a nuclease can hold a nucleotide-like molecule in a catalytic geometry [2].
Serratia marcescens nuclease provides another important mechanistic reference point. Its three-dimensional structure at 1.7 Å resolution helped explain how a nonspecific nuclease can attack nucleic-acid substrates without requiring a long, exact sequence match [3]. That broad activity is the mechanistic basis for why Serratia-type nuclease systems are often discussed in the context of DNA/RNA reduction rather than sequence-specific editing.
BAL 31 nuclease illustrates exonuclease behavior. Mechanistic studies described how BAL 31 acts from nucleic-acid ends and how its action differs from a purely internal-cutting enzyme [4]. Kinetic work on BAL 31 with small substrates and single-stranded DNA further showed that substrate size and structure influence how nuclease digestion proceeds [8].
DNA repair nucleases show the other end of the specificity spectrum. UvrABC excision nuclease recognizes DNA damage and makes cuts around lesions such as psoralen crosslinks, converting a damaged DNA region into a removable segment [9]. FAN1 is another repair nuclease involved in processing DNA interstrand crosslinks, where the enzyme must act on a structurally abnormal DNA substrate rather than simply degrade all DNA present [10].
These examples show why nuclease should be understood as a functional enzyme class. The shared chemistry is phosphodiester bond cleavage, but the biological logic can range from broad degradation to precise lesion processing.
Nucleases are not only industrial tools; they are biological control enzymes. In DNA repair, nucleases remove damaged or inappropriate DNA structures so that repair synthesis and ligation can restore genome integrity. The UvrABC system, for example, has been studied as a model for how cells recognize damaged DNA and make incision events around the lesion [1].
Other nucleases act in immunity or competition. Nuclease colicins are antibacterial toxins that enter susceptible Escherichia coli cells and damage essential nucleic-acid targets, while producing organisms use immunity proteins to protect themselves [11]. This demonstrates a different nuclease function: targeted biological attack rather than process cleanup.
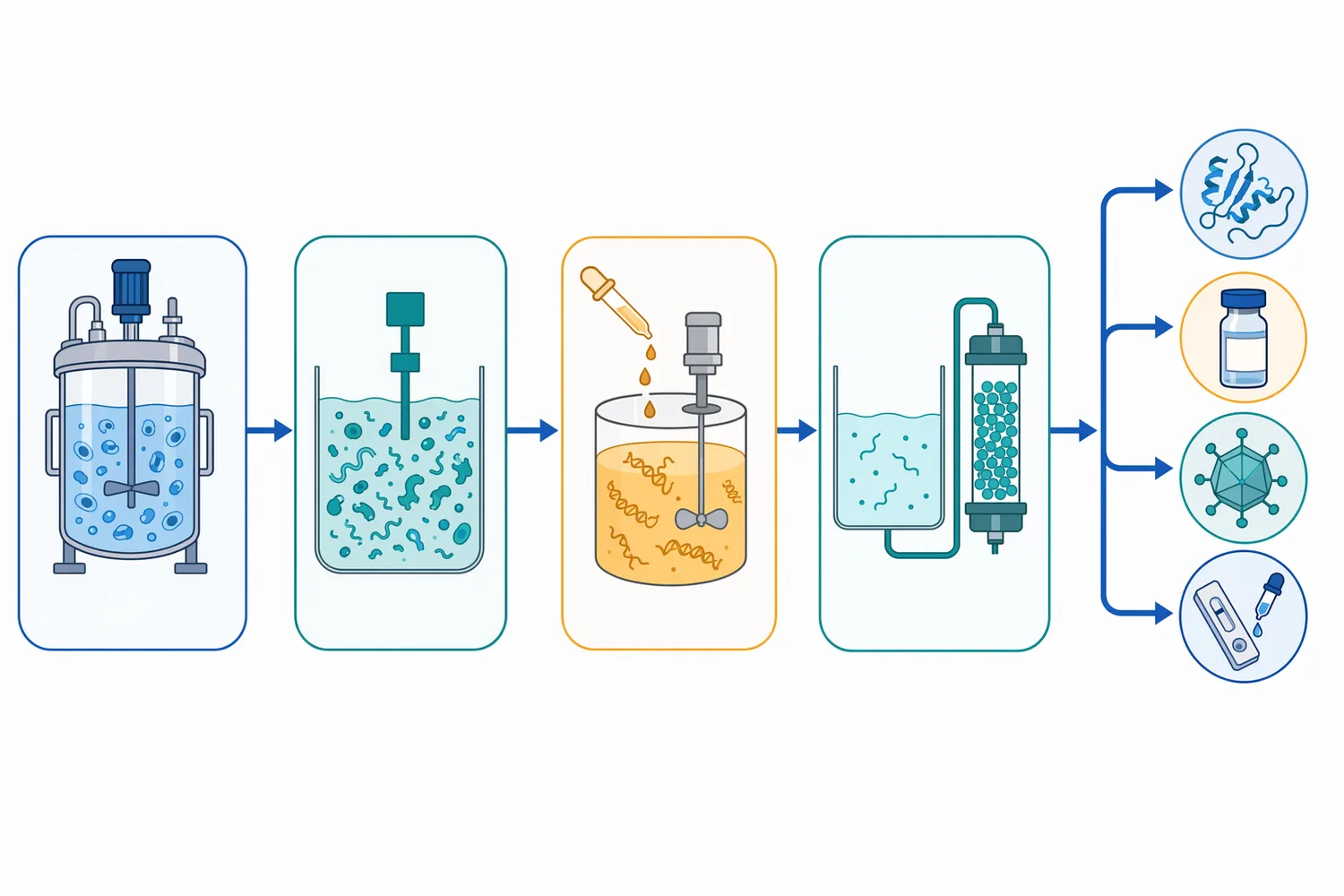
Viral and pathogen-associated nucleases are also active research areas. SARS-CoV-2 nsp15 is an endoribonuclease, and mechanistic studies have considered it as a therapeutic target because viral RNA processing can contribute to infection biology [12]. These systems are not the same as a general commercial nuclease for cleanup, but they illustrate how central nuclease chemistry is across biological systems.
RNA-directed regulation also involves nuclease logic. In RNA interference, small interfering RNA guides gene silencing pathways that ultimately reduce target RNA, linking nucleic-acid recognition with enzymatic cleavage or degradation processes [13]. This is another example of how biological systems use nucleic-acid sequence information to direct nuclease-related outcomes.
Because genome editing is widely known, many buyers and technical readers associate the word nuclease with CRISPR, zinc finger nuclease, or transcription activator-like effector nuclease systems. Those are real nuclease technologies, but they are different from general-purpose nuclease products used for bulk DNA/RNA degradation.
Programmable nucleases are designed to cut a chosen DNA sequence. Zinc finger nuclease systems use engineered DNA-binding protein domains. TALE nuclease systems use transcription activator-like effector DNA-binding domains linked to a cleavage domain. CRISPR-associated nucleases use guide nucleic acids or related targeting logic to direct cleavage. Their purpose is to introduce a site-specific break in a genome, not to reduce all nucleic acid in a process stream.
General nuclease, by contrast, is valued when the goal is broad nucleic-acid breakdown. The desired outcome is not a precise edit but a reduction in intact DNA or RNA length. This difference is important for expectations: a cleanup nuclease should be understood as a processing enzyme, while a programmable nuclease is a genome-engineering tool.
Emerging systems further expand the nuclease landscape. DNA-targeting short Argonautes have been studied in complexes with effector proteins that produce collateral nuclease activity and bacterial population immunity [14]. These advanced systems reinforce the same principle: nuclease chemistry can be nonspecific, structure-specific, damage-specific, or sequence-guided depending on the enzyme system.
Single-stranded nucleic acids often behave differently from double-stranded nucleic acids. Regions of single-stranded DNA or RNA can appear during replication, transcription, damage processing, denaturation, or hybridization workflows. Enzymes such as S1 nuclease—also searched as nuclease S1—are associated with cleavage of single-stranded nucleic-acid regions, making them conceptually different from broad nucleases used simply to reduce bulk DNA/RNA burden.
This distinction matters because substrate structure changes enzyme access. In double-stranded DNA, bases are paired and the backbone follows a helical geometry; in single-stranded regions, bases and backbone can be more exposed and flexible. A nuclease that prefers one structure may not behave the same way on the other. Mechanistic studies of nucleases such as BAL 31 on single-stranded DNA show that single-stranded substrate form can meaningfully affect digestion behavior [8].
For process cleanup, the substrate is often a mixture of nucleic-acid forms: genomic DNA, sheared DNA, RNA, chromatin-associated DNA, and partially denatured regions. A broad nuclease can be useful when multiple nucleic-acid forms are present, while structure-specific nucleases are typically used for more specialized molecular biology purposes.
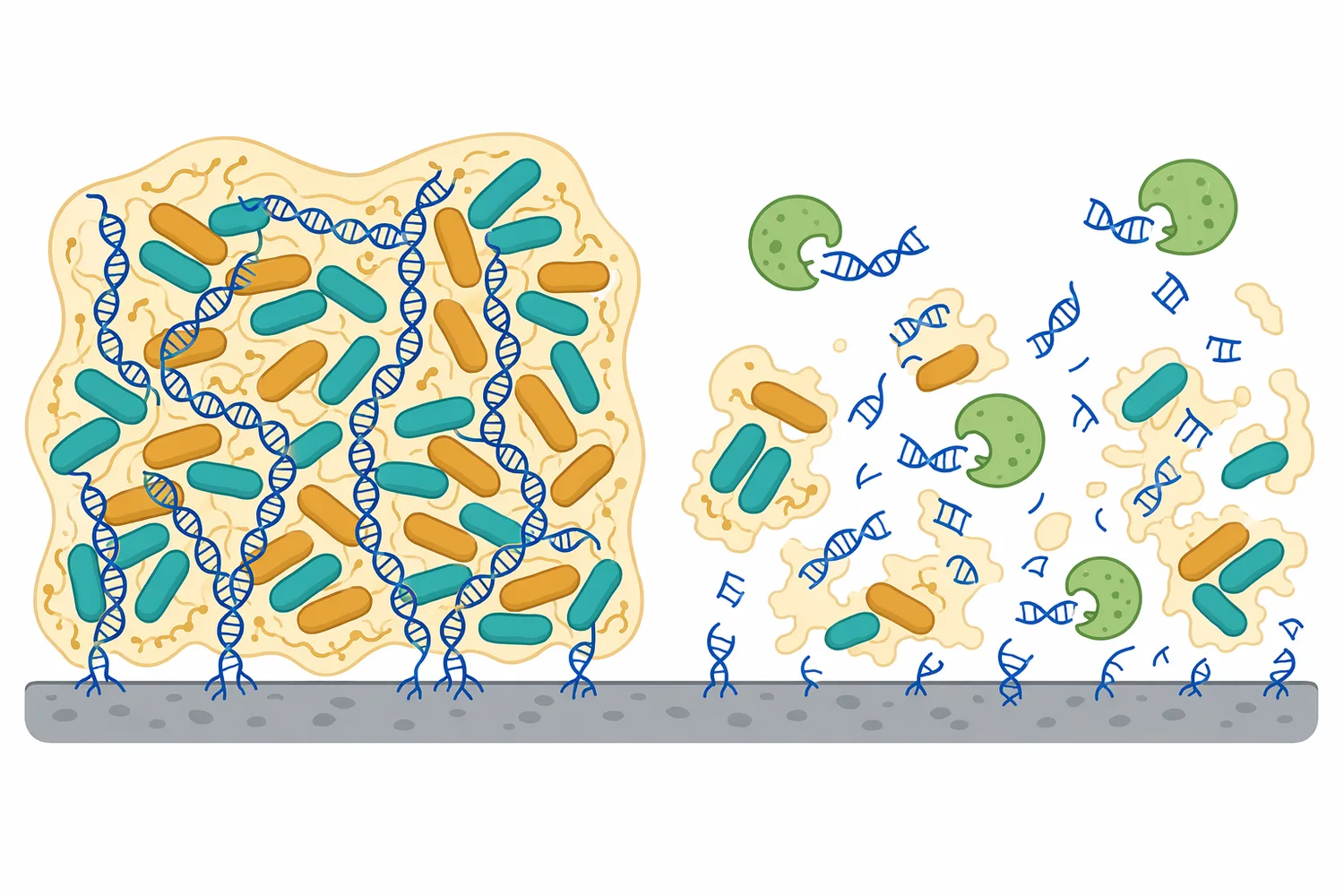
Extracellular DNA can be an important structural component in some microbial biofilms. In those systems, DNA is not only genetic material; it can help form or stabilize the extracellular matrix that holds cells together on a surface. A nuclease can change that matrix by cutting extracellular DNA, weakening one of the polymeric components that contributes to biofilm structure.
The mechanism is again physical as well as chemical. When extracellular DNA strands remain long, they can connect matrix components and contribute to surface-associated structure. When nuclease cuts those strands, the matrix loses part of its polymeric continuity. This does not mean nuclease is automatically a stand-alone antimicrobial; it means nuclease can be relevant where extracellular DNA is a meaningful structural element.
Nuclease-related biofilm work should therefore be viewed as context-dependent. Different organisms and surfaces produce different matrices, and not every biofilm relies on DNA to the same extent. Nuclease colicin and bacterial immunity literature also shows that nuclease activity in microbial systems can have many different biological roles, from matrix disruption to direct nucleic-acid toxicity [11].
Technical users often encounter two opposite nuclease concepts. In one workflow, nuclease is intentionally added to degrade DNA or RNA. In the other, nuclease contamination must be avoided because the goal is to preserve intact nucleic acids. That is why phrases such as nuclease-free water, water nuclease free, and nuclease free water appear frequently in molecular biology purchasing and protocol language.
Nuclease-free water is not a nuclease product; it is water prepared or supplied for workflows where unwanted nuclease activity could damage DNA or RNA. By contrast, a nuclease enzyme product is intentionally used when degradation is desired. Confusing these two concepts can lead to the wrong expectation: one protects nucleic acids, the other breaks them down.
In DNA/RNA cleanup, the presence of active nuclease is the useful feature. In PCR setup, RNA handling, sequencing preparation, or nucleic-acid storage, unwanted nuclease exposure can be damaging. The correct interpretation depends entirely on whether the workflow’s goal is nucleic-acid preservation or nucleic-acid removal.
After nuclease treatment, the most important change is molecular size. Long DNA or RNA strands are converted into shorter fragments. This can reduce viscosity caused by intact genomic DNA, decrease the persistence of long charged polymers, and make nucleic-acid material more compatible with downstream removal.
The second change is accessibility. A large DNA molecule may be entangled with proteins, chromatin material, or other cellular debris. Cutting the backbone creates smaller pieces that can redistribute and separate more readily. In repair biology, nucleases such as UvrABC create incision events that convert damaged DNA into a processable repair intermediate; in industrial cleanup, the same broad principle applies in a nonspecific way: cleavage turns a difficult macromolecule into smaller material [9].
The third change is functional. Intact RNA may encode information, participate in complexes, or interfere with analysis. RNase-type nuclease activity can remove or reduce RNA, while DNase-type activity can reduce DNA. In RNA biology, nuclease-linked pathways are central to controlling whether RNA molecules persist or are destroyed [13].
A nuclease is therefore best understood as a molecular conversion tool. It changes nucleic acids from long, intact polymers into shorter hydrolysis products. Any process benefit—easier handling, lower nucleic-acid burden, improved cleanup—flows from that chemical conversion.
Enzymes.bio supplies Nuclease directly online by the 1 kg unit. The purchasing model is simple: the buyer places the order online, pays online, and the order is processed and shipped. A Certificate of Analysis and Safety Data Sheet are provided with the order for responsible documentation and handling.
This article is intended to support informed use of the enzyme category without implying that one nuclease behaves like every other nuclease. The scientific literature shows a wide range of nuclease mechanisms—from micrococcal nuclease degradation of DNA, to BAL 31 exonuclease action, to UvrABC and FAN1 repair systems, to programmable nuclease technologies—so the word “nuclease” should always be read in the context of the intended application [5].
For buyers who need broad DNA/RNA breakdown, nuclease is relevant because it attacks the chemical backbone of nucleic acids. For workflows that must protect DNA or RNA, nuclease exposure is something to avoid. That contrast is the practical meaning behind both sides of the terminology: nuclease for degradation, nuclease-free water for preservation.
Nuclease is an enzyme class that breaks DNA and/or RNA by hydrolyzing phosphodiester bonds. This cleavage converts long nucleic-acid polymers into shorter fragments, which can reduce nucleic-acid burden, improve cleanup behavior, and support workflows where DNA or RNA must be removed or controlled.
The evidence base is broad. Structural studies at 1.5 Å and 1.7 Å resolution have shown how nuclease active sites organize substrate and metal-assisted chemistry; mechanistic studies of BAL 31, micrococcal nuclease, UvrABC, FAN1, and other systems show that nuclease activity can be broad, end-directed, damage-specific, or biologically targeted depending on the enzyme [2].
Enzymes.bio offers Nuclease for direct online purchase in 1 kg units, with order processing and shipping after online payment. Each order includes a Certificate of Analysis and Safety Data Sheet.
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Nuclease →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.