Pectin methylesterase(PME)는 펙틴의 메틸에스터기를 제거해 자유 카복실기를 늘리고, 그 결과 펙틴의 전하, 칼슘 결합성, 겔화 성향, 세포벽 물성을 바꾸는 효소입니다. 식품 공정에서는 과일·채소 원료의 펙틴 거동을 조절하고, 주스 안정성·청징·텍스처 설계·저메톡실 펙틴 기능화와 연결해 검토됩니다. Enzymes.bio는 PME를 제조하거나 시험하는 실험실이 아니라, 1kg 단위로 온라인 직접 판매하는 효소 공급업체이며 주문 시 CoA와 SDS가 함께 제공됩니다.
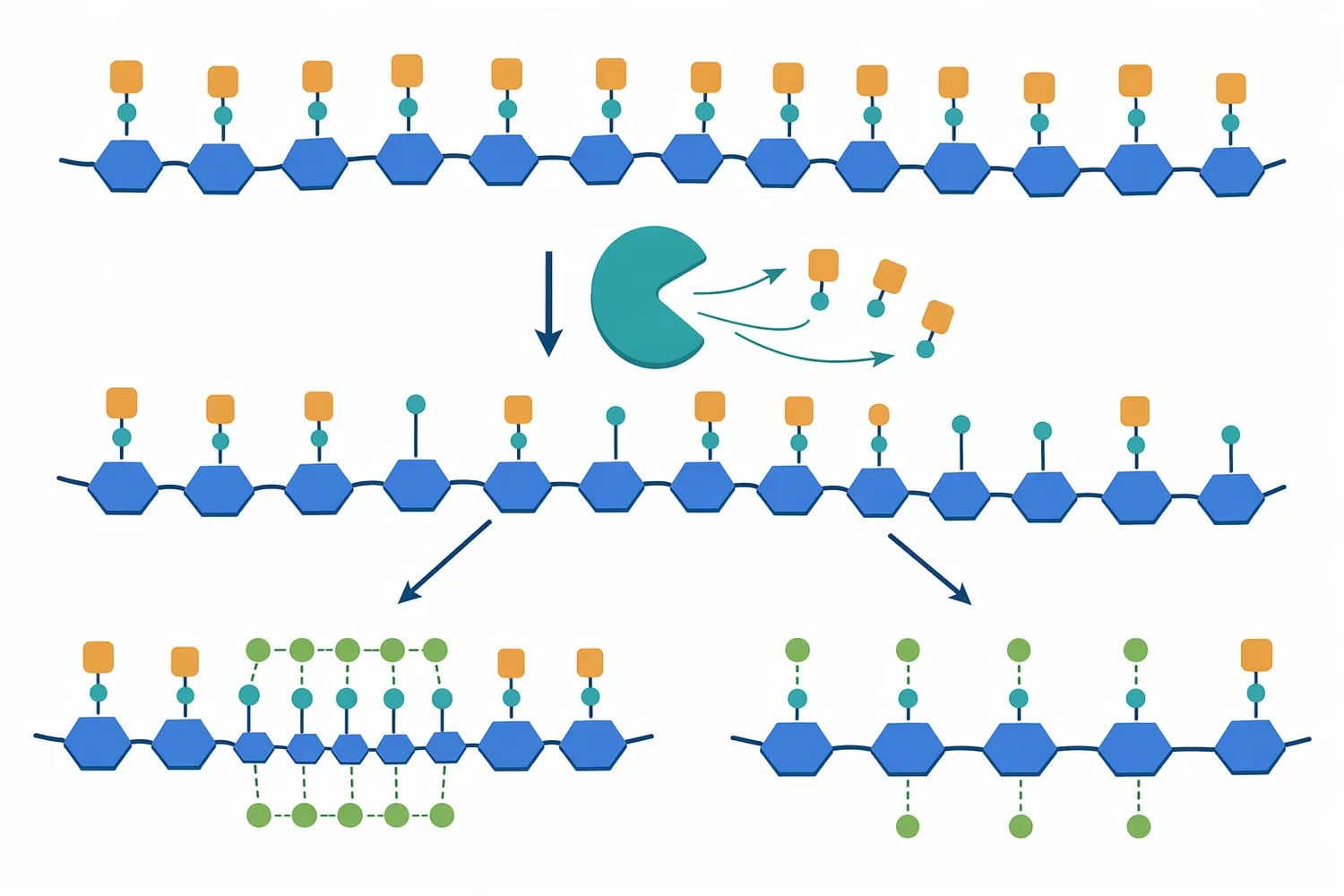
Pectin methylesterase, 흔히 PME 또는 펙틴 메틸에스터레이스라고 부르는 효소는 식물 세포벽의 대표적인 구조 다당류인 펙틴에서 메틸에스터화된 갈락투론산 잔기의 에스터 결합을 가수분해합니다. 이 반응은 펙틴 주사슬을 바로 절단하는 것이 아니라, 펙틴 표면의 화학적 상태를 바꾸는 반응입니다. 따라서 PME의 기능은 “펙틴을 분해한다”기보다 “펙틴이 칼슘, 물, 다른 효소, 세포벽 성분과 상호작용하는 방식을 바꾼다”고 설명하는 편이 정확합니다. 토마토 PME·PME inhibitor 통합 분석에서도 PME는 펙틴 탈메틸에스터화와 조직별 세포벽 조절에 관여하는 효소군으로 다뤄집니다[1].
펙틴은 과일 껍질, 베리류·과즙 부산물, 채소 조직, 식물성 가공 부산물에 풍부하며, 원료별로 분자량, 에스터화 정도, 중성당 가지 구조, 회분·폴리페놀 동반 성분이 달라집니다. 과일 및 베리 주스 생산 부산물에서 얻은 펙틴 연구는 원료에 따라 물리화학적 특성, 항산화성, 유변 특성이 달라질 수 있음을 보여주며, 이는 PME 처리 결과가 “펙틴 함량”만이 아니라 펙틴 구조 전체에 의해 좌우된다는 점을 뒷받침합니다[2].
식품·식물성 원료 가공에서 pectin methylesterase in food가 중요한 이유는, 펙틴이 점도·탁도·침전·겔화·조직감에 직접 영향을 주기 때문입니다. 주스에서는 펙틴이 안정한 cloud를 유지하기도 하고, 반대로 여과와 청징을 어렵게 하기도 합니다. 과일 조직에서는 펙틴의 탈메틸에스터화가 칼슘 가교를 증가시켜 단단함을 높일 수 있지만, 다른 펙틴분해효소가 뒤따르면 연화가 가속될 수도 있습니다. 구기자 숙성 연구에서도 PME와 PME inhibitor가 펙틴 탈메틸화와 탈중합을 조절해 과실 텍스처 특성에 관여하는 주요 조절자로 제시되었습니다[3].
PME 반응의 직접 결과는 펙틴 내 자유 카복실기 증가입니다. 메틸에스터화된 갈락투론산은 상대적으로 전하가 가려져 있지만, PME가 메틸기를 제거하면 음전하를 띠는 카복실기가 드러납니다. 이 변화는 펙틴 사슬 사이의 정전기적 반발, 칼슘 이온과의 결합, 물 결합, 다른 세포벽 고분자와의 상호작용을 바꿉니다. 양파 표피 세포벽 연구에서는 PME 처리가 세포벽을 선택적으로 연화시키면서도 산 유도 creep은 줄이는 복합적인 물성 변화를 보였고, 이는 PME가 단순 연화 효소가 아니라 세포벽 기계적 반응을 재배열하는 효소임을 보여줍니다[4].
PME 처리 후 칼슘이 충분히 존재하면, 인접한 탈메틸화 갈락투론산 구간이 calcium pectate 형태의 가교 네트워크를 만들 수 있습니다. 이 구조는 저메톡실 펙틴 겔의 기본 원리와 연결됩니다. 당을 많이 넣어야 겔이 형성되는 고메톡실 펙틴과 달리, 저메톡실 펙틴은 칼슘과의 이온성 가교를 통해 겔을 형성할 수 있기 때문에 저당 식품, 식물성 겔, 기능성 소재 설계에서 중요합니다. 당근 열안정성 연구에서도 아세트산과 염화칼슘의 상승효과가 펙틴 가교와 세포벽 강화 메커니즘으로 설명되어, 탈메틸화 펙틴과 칼슘 결합의 구조적 의미를 잘 보여줍니다[5].

다만 PME가 항상 단단함을 높이는 방향으로만 작동하는 것은 아닙니다. PME가 펙틴에 자유 카복실기를 만들면 칼슘 가교가 생길 수도 있지만, 동시에 polygalacturonase 같은 효소가 더 쉽게 작용할 수 있는 기질 상태를 만들 수도 있습니다. 즉, 같은 PME라도 공정 내 칼슘 농도, pH, 온도, 원료 펙틴의 메틸에스터화 패턴, 다른 pectinase 존재 여부에 따라 결과가 “겔화”, “조직 강화”, “연화”, “청징 보조”로 달라질 수 있습니다. 사과 연구에서 MdMYB44가 pectin methylesterase 유전자 MdMPE3 발현을 직접 활성화해 과실 crispness를 양의 방향으로 조절한 결과는, PME 조절이 과실 조직감 형성에 구체적으로 연결될 수 있음을 보여줍니다[6].
펙틴 가공에서 PME를 이해하려면 다른 펙틴 관련 효소와 구분해야 합니다. PME는 에스터기를 제거하지만, polygalacturonase는 탈메틸화된 갈락투론산 사슬을 절단하고, pectin lyase는 메틸에스터화된 펙틴을 절단할 수 있습니다. PME inhibitor(PMEI)는 반대로 PME 활성을 억제해 펙틴 탈메틸화의 위치와 정도를 제한합니다. 과일 주스용 pectinase 리뷰는 주스 추출·청징·수율 개선에서 여러 펙틴분해효소가 조합적으로 다뤄진다는 점을 정리하며, PME를 전체 pectinase 네트워크의 일부로 보아야 함을 시사합니다[7].
| 구분 | 직접 작용 대상 | 주된 구조 변화 | 공정상 의미 | 주의할 점 |
|---|---|---|---|---|
| Pectin methylesterase(PME) | 메틸에스터화된 갈락투론산 잔기 | 메틸에스터기 제거, 자유 카복실기 증가 | 칼슘 반응성, 겔화, 세포벽 물성, 후속 효소 접근성 조절 | 주사슬을 직접 절단하지 않으므로 단독 청징 효과는 조건 의존적 |
| Polygalacturonase | 주로 탈메틸화된 homogalacturonan | 갈락투론산 주사슬 절단 | 점도 저하, 펙틴 탈중합, 청징 보조 | PME 처리 후 더 잘 작동할 수 있음 |
| Pectin lyase / pectate lyase | 메틸화 또는 탈메틸화 펙틴 구간 | β-elimination 방식의 사슬 절단 | 빠른 펙틴 분해, 주스·추출 공정 보조 | 원료 펙틴의 에스터화 상태에 따라 효율 변화 |
| PME inhibitor(PMEI) | PME 단백질 | PME 작용 억제 | 과실 조직감, 균열, 세포벽 안정성 조절 연구에 중요 | 효소 처리제가 아니라 조절 단백질로 이해해야 함 |
PME inhibitor는 산업 효소 사용자가 직접적으로 찾는 효소 제품은 아닐 수 있지만, pectin methylesterase inhibitor 연구는 PME 기능을 해석하는 데 중요합니다. 벼 PME inhibitor 연구는 OsPMEI 단백질들이 식물 PME 활성을 억제할 수 있음을 분자·생화학적으로 특성화했고, 이는 식물체가 PME를 단순히 많이 만들기보다 억제자를 통해 공간적·시간적으로 조절한다는 점을 보여줍니다[8]. 키위 PMEI 연구에서도 pH가 inhibitor 구조와 기능에 영향을 줄 수 있어, PME 억제 역시 공정 조건과 단백질 안정성에 따라 달라질 수 있음을 시사합니다[9].
토마토에서는 PME와 PMEI의 균형이 과실 물성 및 균열 저항성과 연결됩니다. SlPMEI27 의존적 펙틴 변형이 토마토 과실 cracking의 유전적 제어와 관련된다는 연구는, pectin methylesterase activity를 무조건 높이는 것이 항상 바람직한 것이 아니라 목적 조직감에 맞게 조절되어야 함을 보여줍니다[10]. 따라서 pectin methylesterase function을 논할 때에는 “활성 증가=개선”이라는 단순 구도보다, 어느 원료에서 어떤 펙틴 패턴을 만들 것인지가 핵심입니다.
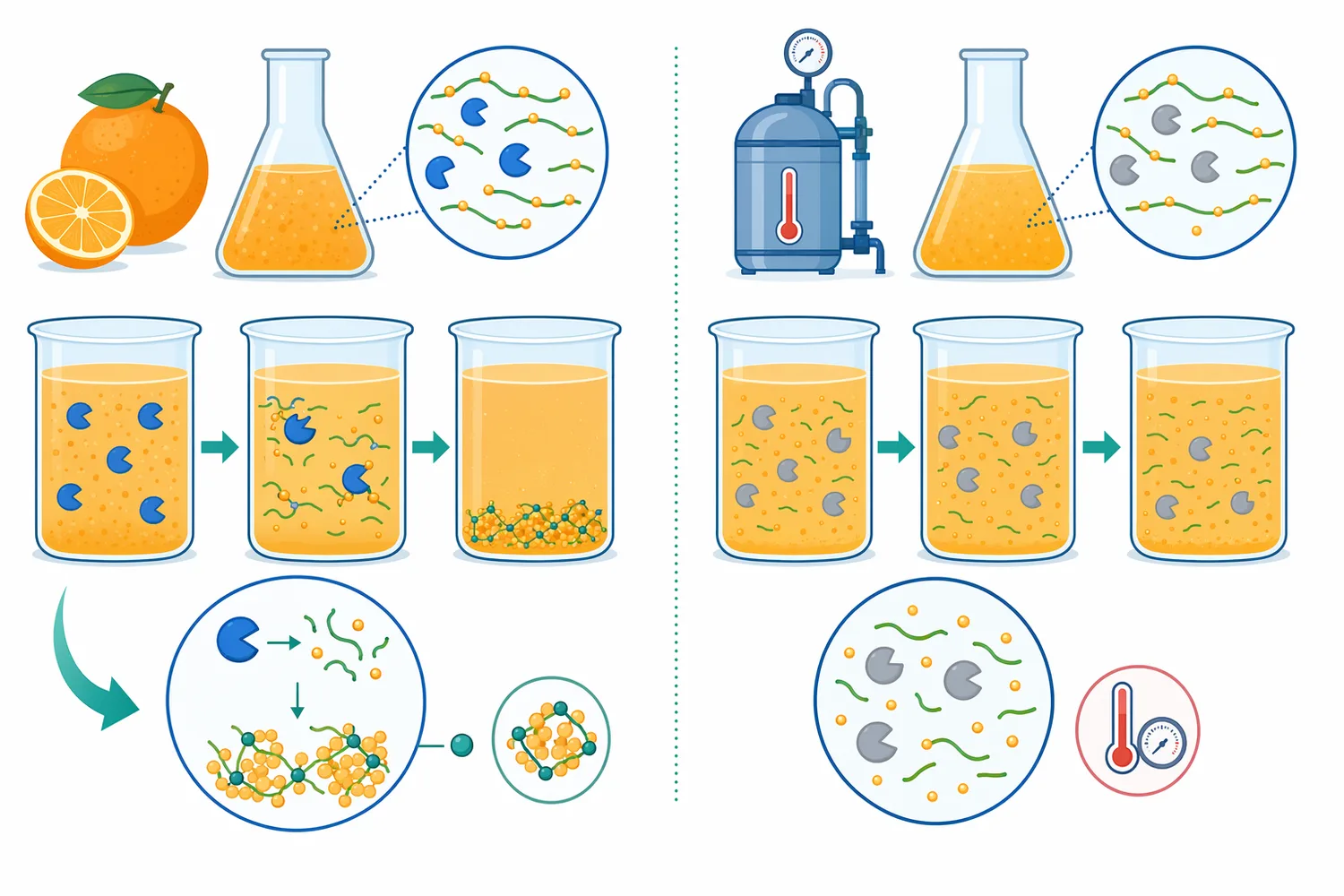
Pectin methylesterase in orange juice는 식품공학 문헌에서 자주 등장하는 주제입니다. 오렌지·만다린 계열 주스에서는 펙틴이 cloud 안정성과 점도에 관여하며, PME가 펙틴을 탈메틸화하면 칼슘과 결합해 불용성 calcium pectate가 형성될 수 있습니다. 이는 경우에 따라 cloud loss, 침전, 상분리로 이어질 수 있어, 주스 품질에서는 PME를 “활용할 것인가”뿐 아니라 “불활성화할 것인가”도 중요한 공정 질문이 됩니다. Nagpur mandarin juice 연구는 열초음파 처리가 PME 불활성화와 cloud stability에 미치는 영향을 다뤄, 감귤 주스에서 PME가 안정성 관리의 핵심 효소임을 보여줍니다[11].
오렌지 자체의 PME 유전자군도 주스 품질과 과육 조직 변화 연구에서 주목됩니다. 네이블 오렌지 연구는 pectin methylesterase gene family를 특성화하고 juice sac granulation과의 관련 가능성을 제시했습니다[12]. 이는 상업적 주스 공정에서 관찰되는 펙틴 문제의 배경에, 원료 과실이 가진 내재 PME·PMEI 시스템과 세포벽 발달 상태가 존재한다는 점을 의미합니다. 따라서 pectin methylesterase in orange juice라는 검색어는 단순히 첨가 효소를 찾는 의미뿐 아니라, 원료 유래 내생 PME의 영향을 이해하려는 맥락에서도 사용됩니다.
비열처리 기술에서도 PME는 품질 지표로 자주 다뤄집니다. Citrus sinensis 주스의 dielectric barrier discharge plasma 처리 연구는 PME 불활성화와 ascorbic acid degradation을 함께 평가해, 비열 공정이 효소 안정성과 영양 성분 보존 사이의 균형 문제를 가진다는 점을 보여줍니다[13]. 자부티카바 주스 연구 역시 초음파 처리에 따른 phenolic compounds, volatile profile, polyphenol oxidase, peroxidase, PME 불활성화를 함께 관찰해, 주스 공정에서 PME가 단독 품질 항목이 아니라 색·향·산화효소와 얽힌 품질 네트워크의 일부임을 보여줍니다[14].
Pectin methylesterase in tomato juice라는 검색 의도는 토마토 주스, 퓨레, 소스, 농축액의 점도·펄프 안정성·조직감을 이해하려는 목적과 연결됩니다. 토마토 PME·PMEI 통합 분석은 Solanum lycopersicum에서 PME와 PMEI의 조직 특이적 발현과 생화학적 특성을 다루며, 토마토 원료가 단일한 펙틴 매트릭스가 아니라 조직·성숙도·유전자 발현에 따라 PME 조절 상태가 달라지는 시스템임을 보여줍니다[1].
토마토 과실 균열 연구는 SlPMEI27을 통한 펙틴 변형이 cracking 저항성과 관련된다는 점을 제시합니다[10]. 토마토 주스나 소스 공정에서 이는 두 가지 실무적 의미를 가집니다. 첫째, 원료 과실의 세포벽 상태가 가공 중 점도와 입자 안정성에 영향을 줄 수 있습니다. 둘째, 외부 PME 처리는 원료에 이미 존재하는 내생 PME·PMEI의 배경 위에 추가되는 조절이므로, 토마토 매트릭스에서의 결과는 사과·감귤·베리와 동일하게 예측할 수 없습니다.
과일 주스 청징에서 PME는 polygalacturonase, pectin lyase, cellulase, hemicellulase 등과 함께 검토되는 경우가 많습니다. 파파야 껍질을 활용한 Aspergillus tubingensis 유래 acidic PME 생산 및 주스 청징 적용 연구는, PME가 산성 조건의 과일 주스 매트릭스에서 청징 공정과 연결될 수 있음을 보여주는 사례입니다[15]. 그러나 PME가 펙틴 주사슬을 직접 절단하지 않는다는 점 때문에, 목표가 빠른 점도 저하나 여과성 향상이라면 다른 pectinolytic 효소와의 역할 분담이 중요합니다.

최근 acid-active endopolygalacturonase 연구는 펙틴 탈중합, pectic-oligomer 생산, fruit juice clarification에 endo-polygalacturonase가 직접적인 역할을 할 수 있음을 보여줍니다[16]. 이와 비교하면 PME는 “절단 효소”라기보다 절단 가능성을 바꾸는 “구조 전환 효소”입니다. 이 차이를 이해하면, PME를 주스 공정에 적용할 때 기대치를 과장하지 않고 설계할 수 있습니다.
PME는 고메톡실 펙틴의 메틸에스터화 정도를 낮추는 효소적 도구로 이해할 수 있습니다. 펙틴 추출·개질 연구에서는 산, 알칼리, 유기산, 열, 초음파, 마이크로파 같은 방법이 사용되며, 원료와 처리 조건에 따라 수율·분자량·에스터화도·항산화성·유변 특성이 달라집니다. 석류 껍질 펙틴 추출 및 화학적 개질 연구는 과일 부산물 펙틴이 구조 변형을 통해 기능성을 바꿀 수 있음을 보여주며, PME 기반 개질도 이러한 “펙틴 기능화” 범주에서 이해할 수 있습니다[17].
농식품 부산물에서 펙틴을 얻는 기술은 자원 효율과 순환경제 관점에서 활발히 연구되고 있습니다. 식품 가공 부산물에서 펙틴을 얻기 위한 신흥 기술 리뷰는 부산물 valorization이 환경 부담 완화와 고부가 소재 생산을 연결하는 전략임을 설명합니다[18]. PME는 여기서 추출 자체를 대체하는 효소라기보다, 회수된 펙틴 또는 펙틴이 풍부한 습식 매트릭스의 에스터화 상태를 조절해 겔화·결합성·후속 분해성을 바꾸는 보조적 기능화 수단입니다.
선인장 열매 껍질처럼 특정 부산물에서 추출한 펙틴도 관심을 받고 있습니다. Opuntia ficus-indica peel 펙틴 생산 연구는 유기산 추출과 공정 최적화를 통해 부산물 기반 펙틴을 생산하고 특성화하는 접근을 다룹니다[19]. 같은 원료 계열에서는 prickly pear fruit의 pectin methylesterase activity와 kinetic parameter를 다룬 연구도 있어, 원료 자체가 가진 내생 PME와 외부 PME 처리 가능성을 구분해 보는 것이 중요합니다[20].
PME는 식품 공정 효소이기 전에 식물 세포벽을 조절하는 생물학적 효소입니다. 과실 숙성, 조직 연화, crispness, 균열, juice sac granulation 같은 현상은 모두 펙틴의 메틸에스터화 상태와 관련될 수 있습니다. 사과 연구에서 MdMYB44가 PME 유전자인 MdMPE3 발현을 직접 활성화해 과실 crispness를 긍정적으로 조절했다는 결과는, PME가 과실 조직감의 분자적 조절축에 포함될 수 있음을 보여줍니다[6].

구기자 숙성 연구도 PME와 PMEI가 펙틴 demethylation 및 depolymerization을 조절하며 과실 텍스처 특성의 주요 조절자로 작용한다고 설명합니다[3]. 여기서 중요한 점은 PME와 펙틴 탈중합이 연결되어 있지만 동일한 반응은 아니라는 것입니다. PME가 먼저 탈메틸화 패턴을 바꾸고, 그 후 polygalacturonase 등 다른 효소가 사슬 절단을 일으키면 조직 연화가 진행될 수 있습니다. 반대로 칼슘 가교가 우세한 조건에서는 조직 강화로 관찰될 수 있습니다.
관련 검색어인 pectin methylesterase arabidopsis는 모델식물 Arabidopsis에서 세포벽, 생장, 스트레스 반응, 발달을 연구할 때 자주 사용됩니다. 이 검색 흐름은 PME가 상업적 식품 효소만이 아니라 식물 생리학의 핵심 효소군이라는 점을 보여줍니다. 다만 Enzymes.bio의 제품 문맥에서는 Arabidopsis 연구 결과를 특정 식품 공정 성능으로 곧바로 환산하기보다, PME가 펙틴 구조를 조절하는 보편적 기전을 뒷받침하는 배경으로 이해하는 것이 적절합니다.
PME의 결과는 원료 펙틴의 에스터화도와 분포에 크게 좌우됩니다. 같은 평균 에스터화도라도 메틸기가 사슬을 따라 무작위로 분포하는지, 블록 형태로 남아 있는지에 따라 칼슘 가교 가능성과 polygalacturonase 접근성이 달라집니다. PME는 효소 기원에 따라 탈메틸화 패턴이 달라질 수 있으며, 이 차이가 최종 점도·겔 강도·침전성에 반영됩니다. 토마토 PME·PMEI 연구처럼 여러 isoform과 억제자가 존재하는 식물 시스템에서는 “PME”라는 이름 아래 다양한 단백질 기능이 공존할 수 있습니다[1].
pH도 중요합니다. PMEI 구조와 기능이 pH에 따라 달라질 수 있다는 키위 PMEI 연구는, PME 조절 단백질의 안정성뿐 아니라 PME 기반 공정 전체가 산도 환경에 민감할 수 있음을 시사합니다[9]. 과일 주스는 대체로 산성 매트릭스이지만, 채소 추출물, 펄프, 식물성 단백질 혼합물, 펙틴 소재 슬러리는 서로 다른 pH 환경을 가집니다. 따라서 PME의 효과는 “효소가 있다”는 사실보다, 해당 매트릭스에서 펙틴이 실제로 접근 가능한 상태인지에 의해 결정됩니다.
칼슘과 다른 다가 양이온의 존재도 핵심 변수입니다. PME가 자유 카복실기를 늘려도 칼슘이 부족하면 강한 가교 네트워크가 형성되지 않을 수 있고, 반대로 칼슘이 충분하면 급격한 점도 증가나 침전이 나타날 수 있습니다. 당근 연구에서 칼슘과 산 조건이 펙틴 가교와 세포벽 강화에 관여한 것은, PME 처리 후의 물성 변화가 효소 반응만으로 끝나지 않고 이온 환경에서 완성된다는 점을 잘 보여줍니다[5].

식품 산업에서 PME는 때로는 투입해 활용하는 효소이고, 때로는 원료에 이미 존재해 불활성화해야 하는 효소입니다. 감귤 주스에서 PME가 계속 작동하면 cloud 안정성이 저하될 수 있으므로, 열처리 또는 비열처리 기술로 PME를 제어하려는 연구가 많습니다. Nagpur mandarin juice의 열초음파 연구는 PME 불활성화와 cloud stability를 연결해, 주스 품질 유지에서 PME 관리가 중요함을 보여줍니다[11].
DBD plasma, ultrasound, microwave 같은 비열 또는 복합 처리 기술은 신선한 맛과 영양 성분을 보존하면서 효소 활성을 낮추려는 목적으로 검토됩니다. Citrus sinensis 주스의 DBD plasma 연구는 PME 불활성화와 ascorbic acid degradation을 함께 평가해, 비열처리도 품질 요소 간 trade-off를 고려해야 함을 보여줍니다[13]. 초음파·마이크로파 복합 식품 처리 리뷰 역시 에너지 효율과 품질 보존을 함께 다루며, PME 같은 효소 안정성이 공정 평가의 중요한 축이 될 수 있음을 시사합니다[21].
이 관점은 외부 PME 사용에도 적용됩니다. PME를 의도적으로 사용한 뒤에는 원하는 탈메틸화 수준에서 반응을 멈추거나, 후속 공정에서 더 이상 영향을 주지 않도록 제어해야 할 수 있습니다. 특히 주스나 퓨레처럼 저장 중 물성이 민감하게 변하는 제품에서는 PME의 잔존 작용이 긍정적일 수도, 부정적일 수도 있습니다. 따라서 PME는 “첨가하면 끝나는 효소”가 아니라, 최종 제품의 안정성 목표와 함께 설계해야 하는 효소입니다.
Enzymes.bio의 Pectin Methylesterase는 펙틴의 메틸에스터화 상태를 조절하고자 하는 식품·식물성 원료 가공, 펙틴 소재 개발, 응용 연구 용도에서 검토할 수 있는 효소 원료입니다. Enzymes.bio는 제조사나 분석 실험실이 아니며, 제품은 1kg 단위로 온라인에서 직접 구매하는 방식으로 제공됩니다. 주문 시 CoA와 SDS가 함께 제공되므로, 제품 확인과 안전 취급 문서로 활용할 수 있습니다.
이 제품의 기술적 위치는 명확합니다. PME는 펙틴 주사슬을 직접 빠르게 절단하는 효소가 아니라, 펙틴의 탈메틸에스터화를 통해 칼슘 반응성, 겔화, 조직감, 후속 펙틴분해효소 작용성을 조절하는 효소입니다. 주스 청징에서 빠른 점도 저하가 목표라면 polygalacturonase나 pectin lyase 계열과의 역할 차이를 이해해야 하며, 반대로 저메톡실 펙틴 특성, 칼슘 기반 겔, 과실 조직감 조절, 원료별 펙틴 기능화가 목표라면 PME의 선택적 화학 개질 기능이 더 직접적인 의미를 가집니다[7].
검색어 관점에서 pectin methylesterase sigma는 실험실 시약이나 문헌 재현을 찾는 사용자가 자주 입력하는 표현일 수 있습니다. 그러나 B2B 식품·소재 공정에서는 브랜드명보다 효소의 실제 기능, 원료 매트릭스, pH·이온 조건, 후속 효소 조합, 최종 물성 목표가 더 중요합니다. Enzymes.bio는 제조사처럼 특정 분석법이나 활성 단위 정의를 제시하는 방식이 아니라, 온라인 구매 가능한 효소 원료와 주문 시 제공되는 문서를 통해 공급을 지원합니다.
PME의 기대 효과는 네 가지로 정리할 수 있습니다. 첫째, 고메톡실 펙틴의 에스터화 상태를 낮추어 저메톡실 펙틴 유사 특성을 강화할 수 있습니다. 둘째, 자유 카복실기를 늘려 칼슘 가교와 겔 형성 가능성을 높일 수 있습니다. 셋째, polygalacturonase 같은 후속 효소가 작용하기 쉬운 펙틴 구조를 만들 수 있습니다. 넷째, 과일·채소 조직의 세포벽 물성을 조절해 텍스처, 연화, crispness, 침전 안정성에 영향을 줄 수 있습니다. 이러한 방향성은 과실 숙성, 토마토, 사과, 감귤, 주스 공정 연구에서 반복적으로 관찰됩니다[3].
한계도 뚜렷합니다. PME 단독으로 모든 펙틴 문제를 해결할 수는 없습니다. 펙틴을 실제로 절단해 분자량을 낮추려면 polygalacturonase나 lyase 계열의 작용이 필요할 수 있고, 주스 cloud 안정성에서는 PME가 오히려 침전 위험을 높일 수도 있습니다. 또한 원료에 이미 내생 PME와 PMEI가 존재하면 외부 PME 처리의 순효과가 달라질 수 있습니다. 토마토와 오렌지 연구에서 보듯이 식물 조직마다 PME 유전자군과 억제자, 세포벽 배경이 다르기 때문에, 한 원료에서의 결과를 다른 원료에 그대로 일반화하기 어렵습니다[12].
따라서 PME는 “펙틴 조절용 정밀 효소”로 보는 것이 가장 정확합니다. 과일 주스에서는 청징 보조 또는 내생 PME 관리라는 양면성을 가지며, 식품 겔에서는 칼슘 기반 구조 형성의 전처리 효소가 될 수 있습니다. 펙틴 소재에서는 에스터화도 조절을 통해 기능성 설계를 돕고, 식물 조직 연구에서는 세포벽 물성과 발달을 이해하는 핵심 효소로 작동합니다. Enzymes.bio에서 공급되는 Pectin Methylesterase는 이러한 용도에서 1kg 단위 온라인 구매가 가능한 효소 원료이며, 주문 시 CoA와 SDS가 함께 제공됩니다.
1kg 단위로 판매되며 재고 보유, 즉시 출고됩니다. 온라인 스토어에서 바로 결제하시면 주문을 처리해 드립니다. 모든 주문에는 시험성적서(CoA)와 물질안전보건자료(SDS)가 포함됩니다.
Pectin Methylesterase 구매하기 →최초 인용 순서로 번호를 매겼습니다. 모든 출처는 발행 시점에 접근 가능 여부를 확인한 오픈 액세스 자료이며, 본문의 인용 번호가 이곳으로 연결됩니다.