Direct answer: Pullulanase enzyme liquid is used in starch hydrolysis because it cuts the α-1,6 branch linkages in amylopectin-derived dextrins, converting branched starch fragments into more linear chains. In glucose and maltose syrup production, that debranching step helps amylase, glucoamylase, and maltose-forming enzymes reach more of the substrate, improving conversion of liquefied starch into fermentable and sweetener sugars [1].
Enzymes.bio supplies Pullulanase Enzyme Liquid for Starch Hydrolysis in Glucose and Maltose Syrup Production as a 1 kg product available for direct online purchase. Buyers complete payment online, after which the order is processed and shipped; a Certificate of Analysis and Safety Data Sheet come with the order .
Pullulanase is best understood as a debranching enzyme for starch conversion. Native starch contains two main glucose polymers: amylose, which is mostly linear, and amylopectin, which is highly branched. The main chains of these polymers are built largely from α-1,4 glycosidic linkages, while the branch points in amylopectin are created by α-1,6 linkages. Pullulanase acts on those α-1,6 branch points, especially in pullulan, amylopectin-derived dextrins, and related branched oligosaccharides, which is why it is widely discussed as an industrial enzyme for starch processing and food applications [1].
That specificity matters because most starch-hydrolysis processes are not limited by the presence of starch alone; they are limited by how accessible the starch chains become after cooking, liquefaction, and partial saccharification. Once α-amylase has reduced viscosity by cutting internal α-1,4 bonds, the process mixture still contains branched dextrins. Those branch points interrupt the smooth action of saccharifying enzymes. Pullulanase removes the branch junctions, producing more linear dextrin chains that are easier for partner enzymes to convert into glucose, maltose, and other soluble sugars [2].
In practice, pullulanase does not replace α-amylase, glucoamylase, or maltose-forming enzymes. It complements them. α-Amylase rapidly opens the starch polymer and lowers viscosity; glucoamylase releases glucose from chain ends; maltose-forming systems generate maltose from accessible linear chains. Pullulanase’s role is to expose more usable chain length by removing α-1,6 “stop points” that otherwise remain in the dextrin mixture [3].
Starch syrup production begins with a granular plant polymer and ends with a controlled carbohydrate solution. During gelatinization and liquefaction, heat and α-amylase disrupt starch granules and break long chains into soluble dextrins. This step is essential because it reduces viscosity and makes the substrate pumpable, but it does not fully erase the branched architecture of amylopectin [4].
The remaining branched dextrins are important because enzymes do not cut all glycosidic bonds with equal ease. Glucoamylase, for example, releases glucose progressively from non-reducing ends, and its productive movement along a chain is affected when it encounters or approaches an α-1,6 branch. Maltose-forming enzymes are also affected by branching because they need accessible linear segments to release maltose units efficiently. When branches remain, some carbohydrate stays as higher oligosaccharides or limit dextrins instead of being converted into the desired syrup sugars [5].
Pullulanase changes that substrate geometry. By cleaving α-1,6 linkages, it separates side chains from the branched amylopectin backbone. The result is not simply “smaller starch”; it is starch with fewer branch junctions and more linear chains. Those newly linearized chains present more accessible non-reducing ends and fewer structural interruptions, so partner enzymes can continue hydrolysis more completely [1].
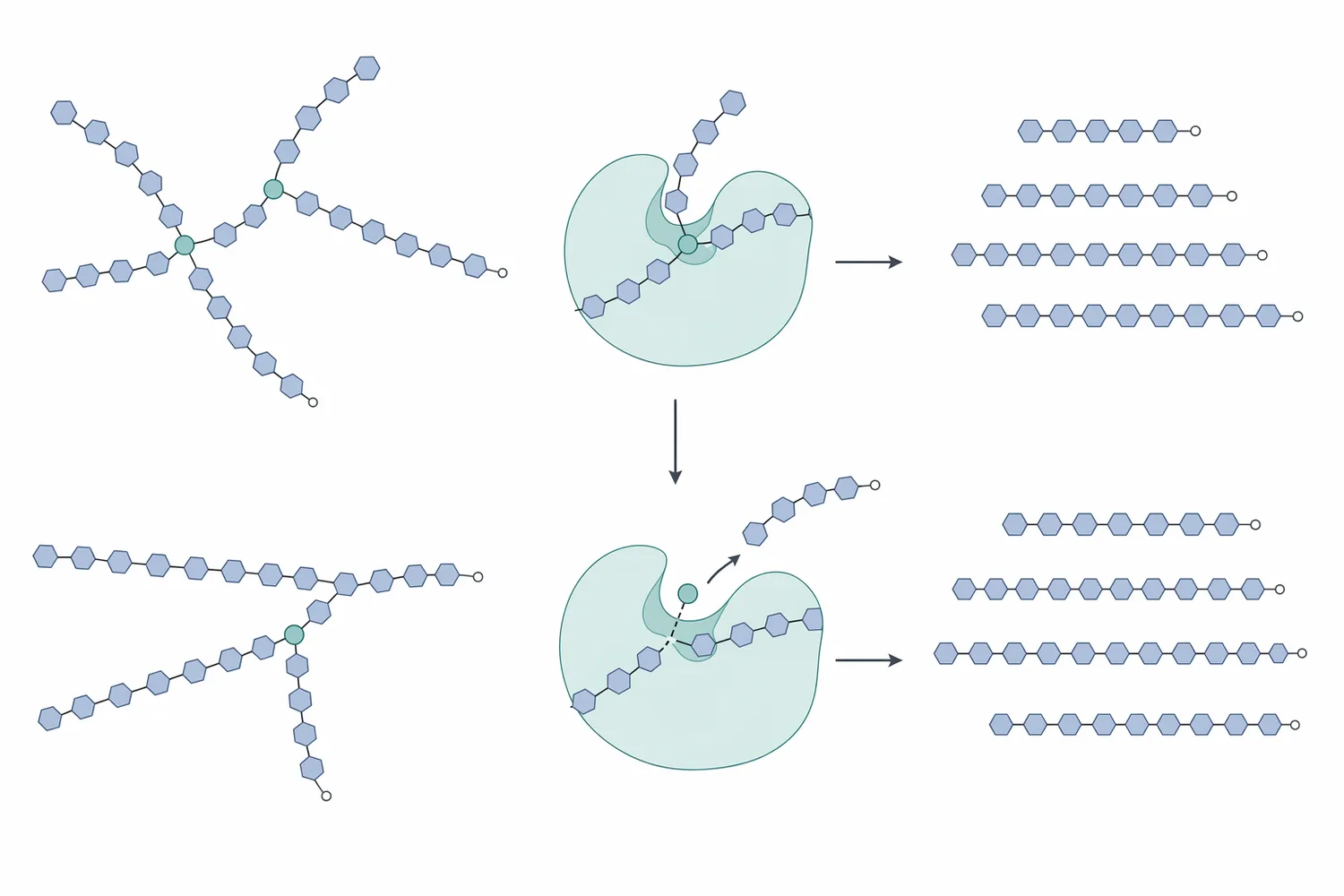
For glucose syrup, this supports deeper conversion because glucoamylase can act on more exposed chain ends and convert dextrins toward glucose. For maltose syrup, debranching helps create longer linear regions that maltose-forming enzymes can use to release maltose rather than leaving branched residues behind. The same chemical event—α-1,6 hydrolysis—therefore supports different syrup profiles depending on which saccharifying enzymes are present [3].
A typical starch-conversion workflow can be viewed as four functional stages: starch preparation, liquefaction, debranching/saccharification, and finishing. Pullulanase is most relevant after starch has been hydrated, heated, and partially broken down, because the enzyme works most effectively when branch linkages are accessible within soluble or swollen starch-derived material rather than locked inside intact granules [6].
During liquefaction, α-amylase attacks internal α-1,4 bonds. This creates a mixture of dextrins with different chain lengths and branching patterns. The process becomes less about reducing bulk viscosity and more about shaping the carbohydrate profile. At this point, pullulanase can be used to remove α-1,6 branches while saccharifying enzymes convert the resulting linear dextrins into the target sugars [4].
The final syrup profile depends on the enzyme system. A glucose-oriented process uses debranching to support glucoamylase-driven glucose release. A maltose-oriented process uses debranching to improve the availability of linear chains for maltose production. A fermentation-feedstock process may value both glucose and maltose because both can contribute fermentable carbohydrate, depending on the organism and downstream process [7].
The important operational concept is that pullulanase improves the structure of the substrate for the next enzymatic step. It does not make every syrup identical. The same debranching action can support high-glucose syrup, maltose syrup, or fermentable sugar streams because the partner enzymes determine how the exposed chains are further hydrolyzed [8].
The starch-hydrolysis enzyme system is easier to understand when each enzyme is viewed by the bond or substrate feature it primarily addresses. Pullulanase occupies a distinct position because it targets the branches that other enzymes often leave behind.
| Enzyme type | Main action in starch conversion | What changes in the substrate | Relevance to glucose or maltose syrup |
|---|---|---|---|
| α-Amylase | Cuts internal α-1,4 linkages in starch chains | Rapidly lowers viscosity and forms soluble dextrins | Prepares liquefied starch for later saccharification |
| Pullulanase | Cuts α-1,6 branch linkages in amylopectin-derived dextrins | Converts branched dextrins into more linear chains | Helps partner enzymes access more carbohydrate for glucose or maltose formation |
| Glucoamylase | Releases glucose from chain ends | Converts accessible dextrins into glucose | Central enzyme for glucose-rich syrups |
| Maltose-forming enzymes | Release maltose from suitable linear dextrin segments | Produces maltose from accessible chain regions | Central to maltose-rich syrup profiles |
| Acid hydrolysis | Non-enzymatic cleavage of glycosidic bonds under acid conditions | Breaks starch less selectively than enzymes | Historically important, but enzymatic processing offers more controlled sugar profiles |
Systematic work comparing enzymatic methods and acid hydrolysis across starch sources highlights why enzymes are favored when the goal is controlled syrup production. Enzymes provide bond-selective reactions, allowing starch processors to target liquefaction, debranching, and saccharification as separate but coordinated steps rather than relying on broad chemical hydrolysis [3].
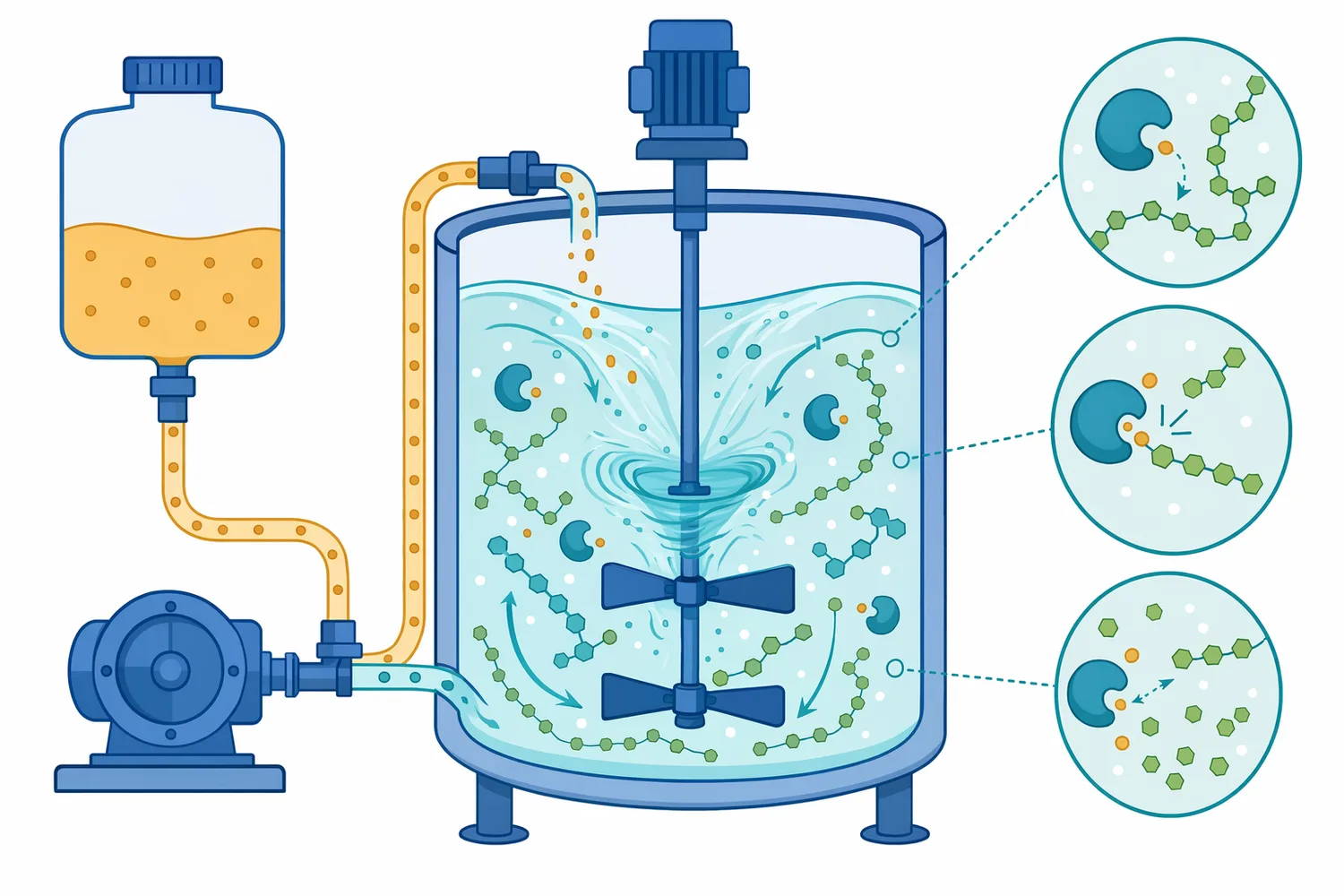
At the molecular level, pullulanase binds a branched starch fragment so that the α-1,6 linkage is positioned in the enzyme’s catalytic site. Enzyme catalysis depends on substrate binding, transition-state stabilization, and precise orientation of reactive groups; this is why the shape of the carbohydrate chain and the accessibility of the branch point directly affect reaction rate [9].
Before pullulanase acts, an amylopectin-derived dextrin may have a main chain with one or more side chains attached through α-1,6 bonds. These side chains create compact, branched structures. Saccharifying enzymes can hydrolyze exposed α-1,4 chains, but their progress slows or stops when the branch point creates steric and chemical constraints. This is the origin of limit dextrins in starch hydrolysis [10].
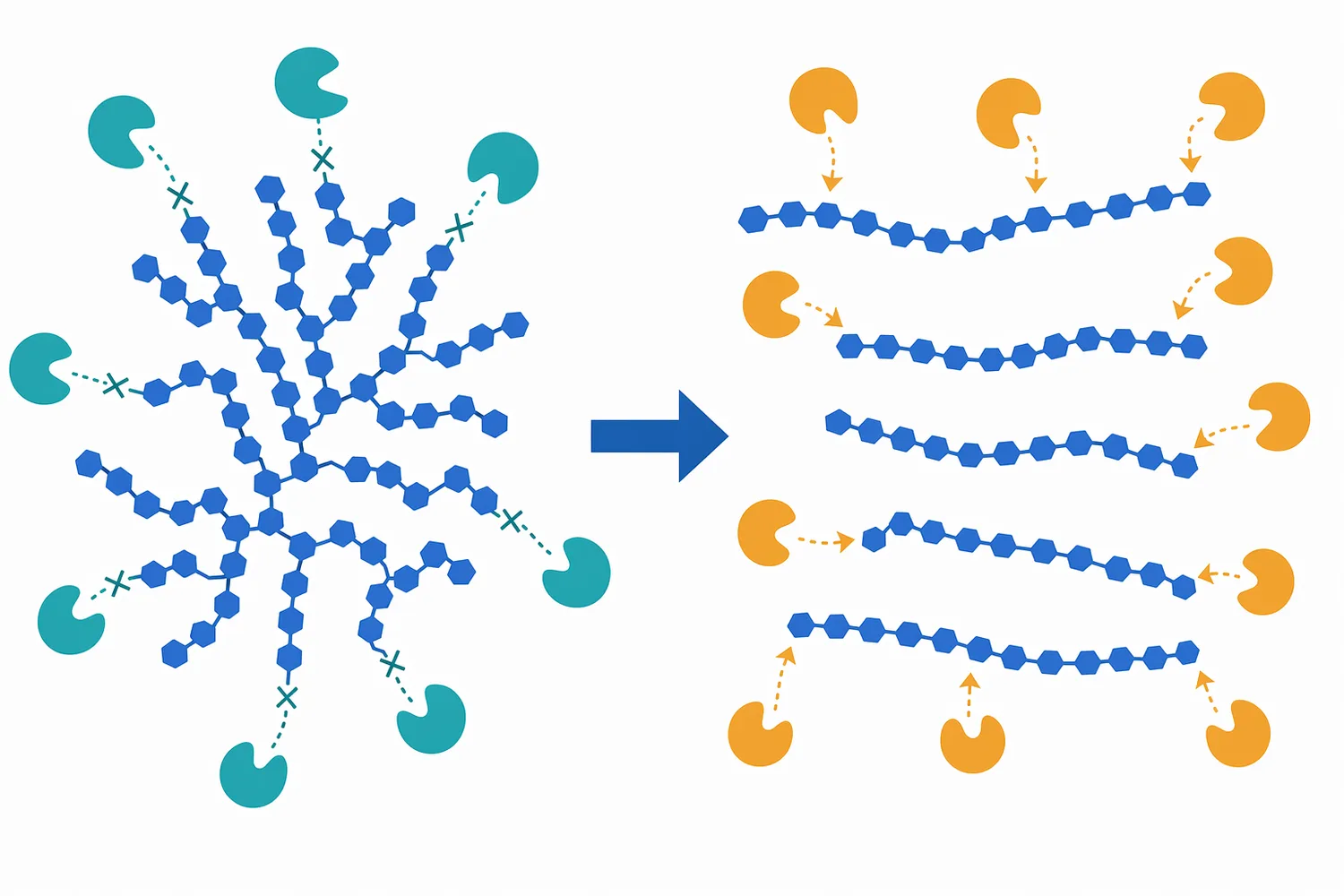
After pullulanase cleaves the branch linkage, the side chain becomes a separate linear α-1,4 glucan. The former branch point is no longer a barrier. The released side chain and the remaining backbone each become better substrates for enzymes that work along linear chains or from non-reducing ends. In glucose production, this increases the number of accessible chain ends for glucose release. In maltose production, it increases the amount of linear chain available for maltose-forming action [1].
This is also why pullulanase can influence conversion without being the enzyme that produces most of the final monosaccharide or disaccharide. Its contribution is structural: it changes the architecture of the dextrin pool. Once the architecture is less branched, the saccharifying enzymes can convert carbohydrate that would otherwise remain partly resistant to the desired extent of hydrolysis [2].
Research on glucose syrup production from different starch sources consistently shows that enzymatic hydrolysis is a central route for converting plant starch into syrup. Studies have examined wheat starch, red sorghum starch, sweet potato starch, cassava starch, and other materials, reinforcing the broader point that starch source and pretreatment influence hydrolysis behavior [11].
A study on glucose syrup production from red sorghum starch specifically examined the effects of substrate and enzyme concentration on syrup production by enzymatic hydrolysis. Although the exact optimal balance depends on the system being studied, the work supports a key processing reality: conversion is governed by the interaction between starch loading, enzyme action, and hydrolysis conditions rather than by the starch source alone [12].
Wheat starch has also been investigated for enzymatic glucose syrup production, showing that conventional cereal starches remain relevant substrates for enzyme-based hydrolysis. For pullulanase users, such studies are important because liquefied wheat starch, corn starch, cassava starch, sorghum starch, and other starches all generate dextrin mixtures in which amylopectin-derived branches can remain after α-amylase treatment [11].
Sweet potato starch syrup production has been evaluated using response surface methodology for scale-up, illustrating that enzyme-based syrup processes are often optimized around interacting variables rather than a single factor. Pullulanase fits this logic because debranching changes the substrate pool available to the rest of the saccharification system, making its effect dependent on the broader process design [13].
Cassava starch research has also examined extrusion and enzymatic hydrolysis in the context of downstream dark fermentative hydrogen production. While that application differs from food syrup production, it reinforces the industrial importance of converting starch into fermentable sugar streams and shows why pretreatment plus enzymatic hydrolysis is a recurring strategy for starch-rich feedstocks [14].
In glucose syrup production, the target is extensive conversion of starch-derived dextrins into glucose. Liquefaction alone cannot achieve that objective because α-amylase mainly shortens chains and lowers viscosity; it does not fully saccharify the material. Glucoamylase then releases glucose from accessible chain ends, but branched dextrins slow complete conversion [5].
Pullulanase supports glucose syrup production by reducing the population of α-1,6-branched dextrins. Once debranched, those chains become more suitable for glucoamylase action. The practical effect is that more of the carbohydrate fraction can move from partially hydrolyzed dextrin into glucose, provided the rest of the saccharification system is aligned with that goal [1].
This mechanism is especially relevant when the process aims for high conversion rather than simply partial hydrolysis. The dextrose-equivalent concept used in syrup production reflects the extent to which starch has been converted into reducing sugars; enzymatic systems can be designed to push conversion by combining liquefaction, debranching, and saccharification rather than relying on a single enzyme function [3].
Different starch sources behave differently because granule structure, amylose-to-amylopectin ratio, prior heat treatment, and residual resistant structures affect enzyme access. Studies on non-conventional starchy materials for glucose syrup production emphasize the need to account for raw-material differences when applying enzymatic conversion strategies [7].
Maltose syrup production has a different objective from glucose syrup production. Instead of pushing all accessible carbohydrate toward glucose, the process is designed to accumulate maltose. That requires enzymes and conditions that favor maltose release from dextrin chains while limiting excessive conversion to glucose [1].
Pullulanase is useful in this context because maltose-forming enzymes need accessible linear chain regions. Branch points interrupt those regions. When pullulanase removes α-1,6 linkages, it creates linear dextrins that can be converted more effectively into maltose by the appropriate saccharifying system [2].
The mechanism is similar to glucose syrup production, but the final sugar outcome is different. In glucose syrup, debranching supports glucoamylase access so glucose can accumulate. In maltose syrup, debranching supports maltose-forming action so maltose can accumulate. Pullulanase therefore helps define the available substrate, while the companion enzymes define the direction of sugar formation [8].
This distinction is important for buyers using a liquid pullulanase product in an established starch process. Pullulanase should be viewed as a way to reduce branch-related limitations in the dextrin pool, not as a standalone determinant of syrup composition. The same debranched substrate can be routed toward different sugar profiles depending on the enzyme system already present [3].
Starch is not a uniform raw material. Granules from different botanical sources vary in size, crystalline organization, amylose content, amylopectin branching patterns, lipid interactions, and resistance to hydration. These features affect how readily enzymes reach glycosidic bonds during hydrolysis [7].
Recent work on pullulanase catalysis across different starch granules emphasizes that the interaction between enzyme and starch surface structure matters. The enzyme must bind productively before it can catalyze cleavage, so the physical state of the starch—granular, gelatinized, liquefied, or partially hydrolyzed—affects how much of the branch structure is actually available for debranching [6].
This is why pullulanase is commonly associated with post-liquefaction or saccharification stages rather than with untreated native starch alone. Gelatinization opens the starch matrix; liquefaction reduces molecular size and viscosity; debranching then addresses the remaining α-1,6 architecture. Each step makes the next step more efficient by changing the physical and chemical form of the substrate [4].
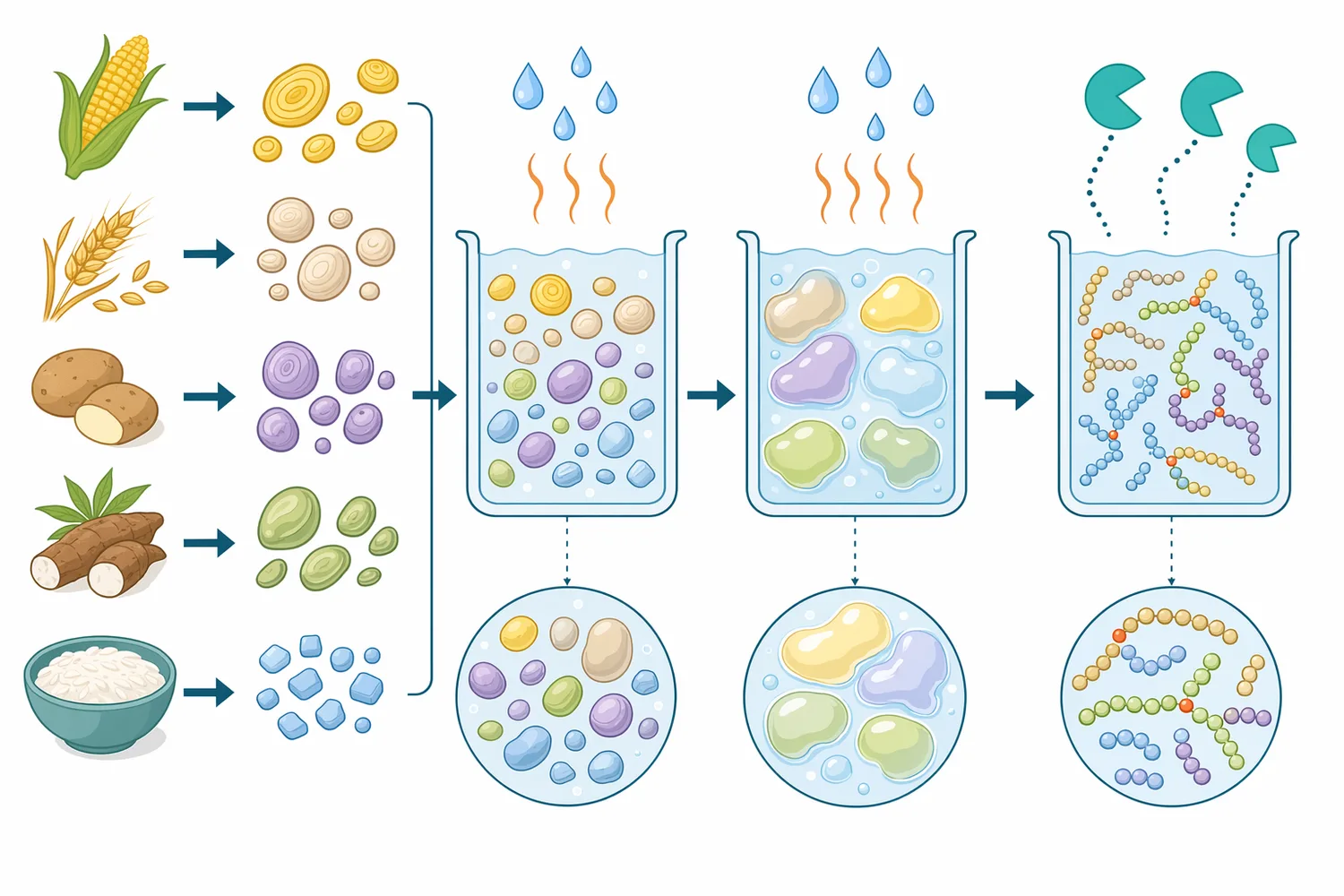
Resistant starch research also helps explain why some starch fractions are more difficult to hydrolyze. Structures such as tightly organized crystalline regions, amylose-lipid complexes, or resistant starch forms can reduce enzyme access even when the correct enzyme is present. Pullulanase removes branch linkages, but it still depends on those linkages being accessible in the processed substrate [10].
Pullulanase is most powerful when used as part of a synergistic enzyme system. Synergy means that one enzyme changes the substrate in a way that improves the action of another. In starch hydrolysis, α-amylase first makes the polymer soluble and manageable; pullulanase removes branch obstacles; glucoamylase or maltose-forming enzymes then convert the exposed chains into target sugars [15].
This sequence is not merely theoretical. Food and starch-processing studies frequently combine amylolytic and debranching activities because starch structure contains both linear and branched features. A single enzyme class rarely addresses all relevant bonds efficiently. By dividing the work across enzymes, the process can be more selective and better matched to the desired carbohydrate profile [16].
The oat milk study using synergistic amylase and debranching enzyme catalysis is not a syrup-production study, but it illustrates the same functional principle: combining amylase action with debranching can alter starch-derived structure and improve product behavior. For syrup production, the analogous benefit is improved carbohydrate accessibility for saccharification [15].
In glucose and maltose syrup processes, this synergy can reduce the amount of branched oligosaccharide left after hydrolysis and support more predictable conversion. The exact syrup composition remains process-dependent, but the biochemical logic of debranching is consistent: fewer α-1,6 barriers means more linear substrate for the enzymes that follow [1].
A liquid enzyme format is convenient for many starch-hydrolysis operations because it can be metered and dispersed into aqueous process streams. In a starch slurry or liquefied dextrin stream, good distribution matters: pullulanase must contact accessible branch linkages throughout the substrate rather than remain localized in one part of the mixture [17].
From a reaction standpoint, enzyme performance is affected by substrate concentration, enzyme concentration, contact time, and the physical accessibility of the substrate. General enzyme-catalysis research shows that reaction rate does not increase indefinitely with substrate because enzyme active sites become saturated; in starch processing, this helps explain why mixing, solids level, and enzyme-substrate contact influence conversion behavior [17].
The liquid format does not change the fundamental mechanism of pullulanase. The enzyme still acts by binding branched dextrins and hydrolyzing α-1,6 linkages. The value of the format is practical handling within an aqueous process: it can be incorporated into the stage where debranching is intended and where the starch has already been made accessible enough for enzymatic action [1].
Enzymes.bio supplies this pullulanase enzyme liquid by the 1 kg unit through direct online purchase. After online payment, the order is processed and shipped, with a Certificate of Analysis and Safety Data Sheet included with the order .
The primary benefit of pullulanase in syrup production is improved use of amylopectin-derived carbohydrate. Amylopectin is highly branched, so a process that does not address α-1,6 linkages can leave more branched dextrin behind. Pullulanase helps convert that branched fraction into linear material that partner enzymes can use [1].
For glucose syrup, the benefit is clearer access for glucoamylase. More linear dextrins and more available chain ends allow glucose release to proceed more completely, assuming the rest of the process supports glucose formation. This is why debranching is often discussed alongside saccharification rather than as a separate, unrelated modification step [5].
For maltose syrup, the benefit is improved availability of linear chain segments. Maltose accumulation depends on enzyme systems that release maltose from dextrins without over-converting the product to glucose. Pullulanase helps by removing the branch points that shorten usable linear regions and create residual limit dextrins [2].
For fermentation feedstocks, the benefit is broader: glucose, maltose, and soluble oligosaccharides can all contribute to fermentable carbohydrate depending on the microorganism and process. Reviews of enzymatic conversion of starchy materials for glucose syrup and related sugar streams show continuing interest in enzyme-based hydrolysis because it can convert diverse starch feedstocks into usable carbohydrate solutions [7].
The strongest and most direct evidence for pullulanase is its established biochemical function as a starch-debranching enzyme. Pullulanase acts on α-1,6 linkages, which explains its role in converting branched amylopectin-derived dextrins into more linear chains. This mechanism is consistently recognized in reviews of pullulanase applications in the food industry [1].
There is also strong practical support for enzymatic starch hydrolysis as a route to glucose syrup production across multiple starch sources. Studies and reviews cover wheat starch, red sorghum starch, cassava starch, sweet potato starch, and non-conventional starchy materials, showing that enzyme-based conversion is a major technical pathway for syrup and fermentable sugar production [3].
What remains process-dependent is the final sugar profile. Pullulanase improves debranching, but glucose concentration, maltose concentration, residual dextrin profile, viscosity, and process time depend on the full enzyme system and operating context. General enzyme kinetics makes this expected: substrate availability, enzyme saturation, and reaction conditions all affect the observed rate and extent of hydrolysis [8].
Adjacent research also supports the broader starch-structuring role of pullulanase. Pullulanase and debranching enzymes are studied not only for syrups but also for starch modification, resistant starch formation, beverage stability, and specialty carbohydrate structures. These applications are not identical to syrup production, but they reinforce the central point that debranching materially changes starch architecture [15].
Pullulanase Enzyme Liquid for Starch Hydrolysis in Glucose and Maltose Syrup Production is intended for buyers who need a debranching enzyme input for starch-conversion processes. Its technical purpose is straightforward: reduce α-1,6 branch barriers in liquefied or otherwise accessible starch-derived dextrins so that the rest of the enzyme system can convert more carbohydrate into glucose, maltose, or fermentable sugars [1].
The product is sold directly online by Enzymes.bio in 1 kg units. Buyers place the order and pay online; the order is then processed and shipped, and the Certificate of Analysis and Safety Data Sheet come with the order .
For starch hydrolysis, the key takeaway is that pullulanase improves the substrate, not just the reaction mixture. By cutting branch points, it turns compact branched dextrins into more linear chains that are easier for glucoamylase and maltose-forming enzymes to process. That concrete structural change is the reason pullulanase is valued in glucose syrup, maltose syrup, and fermentable sugar production [3].
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Pullulanase Enzyme Liquid For Starch Hydrolysis In Glucose And Maltose Syrup Production →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.