إنزيم Pullulanase السائل هو إنزيم نزع تفرّع يُستخدم في تحلل النشا الصناعي لأنه يفتح روابط α-1,6 في الأميلوبكتين والدكسترينات المتفرعة، وبذلك يجعل السلاسل الكربوهيدراتية أكثر قابلية للتحويل إلى جلوكوز أو مالتوز. في إنتاج شراب الجلوكوز يعمل عادةً مع الجلوكوأميليز، وفي إنتاج شراب المالتوز يعمل مع إنزيمات مالتيّة مثل β-amylase أو الأنظمة المالتيّة المناسبة، مع هدف عملي يتمثل في تقليل الدكسترينات المتبقية وتحسين التركيب السكري النهائي [1].
تُورّد Enzymes.bio هذا المنتج كـ Pullulanase Enzyme Liquid For Starch Hydrolysis In Glucose And Maltose Syrup Production للشراء المباشر عبر الإنترنت بوحدة 1 كجم، مع إرفاق CoA و SDS مع الطلب. Enzymes.bio جهة توريد للإنزيمات وليست جهة تصنيع أو مختبرًا، لذلك ينبغي فهم المنتج في سياق استخدام صناعي قائم لا بوصفه وصفة تشغيلية موحدة لكل خط إنتاج .
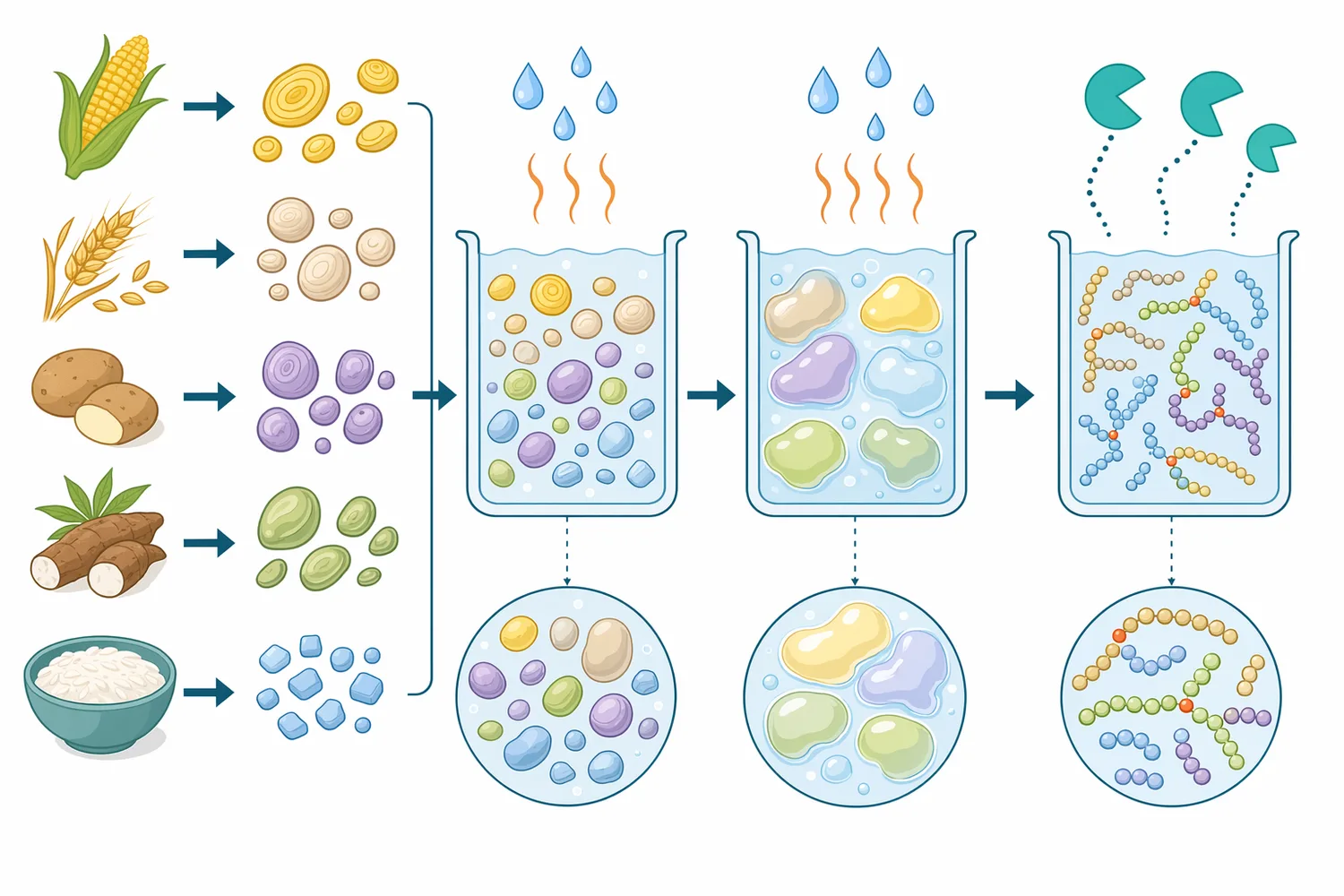
النشا ليس بوليمرًا خطيًا بسيطًا. فهو يتكون أساسًا من أميلوز أكثر خطية بروابط α-1,4، وأميلوبكتين متفرع يحتوي على نقاط تفرّع بروابط α-1,6. عند تحويل النشا إلى شراب جلوكوز أو مالتوز، تُجزّأ السلاسل الطويلة أولًا إلى دكسترينات أقصر، ثم تُستكمل السَكَرة بواسطة إنزيمات متخصصة. المشكلة أن نقاط التفرّع تبقى عائقًا بنيويًا: الإنزيمات التي تعمل على روابط α-1,4 تستطيع التقدم على السلاسل الخطية، لكنها تتباطأ أو تتوقف قرب روابط α-1,6، فتظهر دكسترينات حدّية أو سكريات أعلى غير مرغوبة في الشراب النهائي [2].
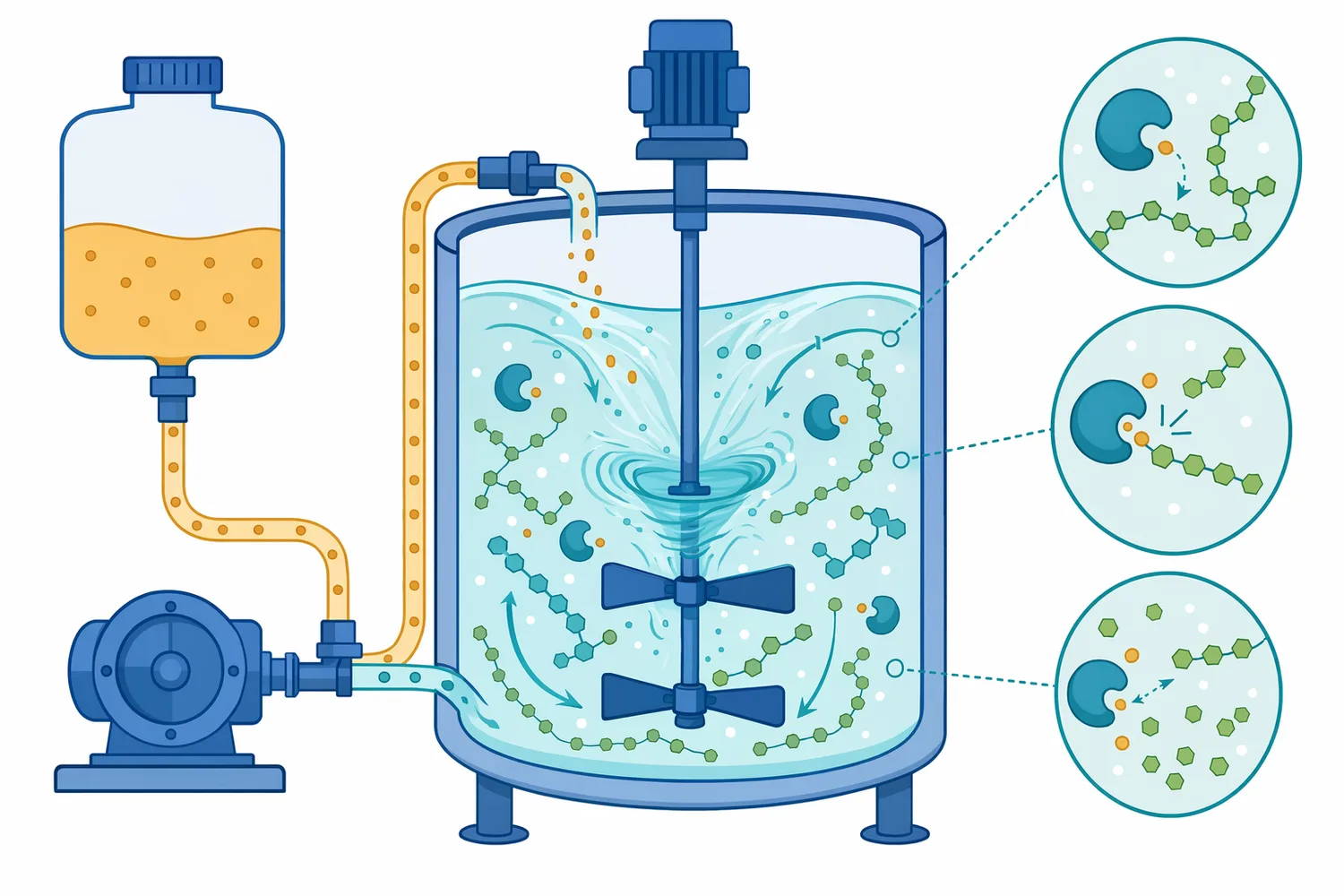
Pullulanase يعالج هذه العقدة البنيوية تحديدًا. فهو لا يُستخدم عادةً باعتباره إنزيمًا وحيدًا يحوّل النشا مباشرةً إلى الجلوكوز أو المالتوز، بل كإنزيم مساعد يزيل التفرعات ويفتح البنية أمام إنزيمات السَكَرة. لذلك تُفهم قيمته الصناعية من خلال التكامل: α-amylase يقلل اللزوجة ويفتح النشا إلى دكسترينات، glucoamylase يحرر الجلوكوز، الإنزيمات المالتيّة تدفع النظام نحو المالتوز، وpullulanase يزيل الحواجز المتفرعة التي تحدّ من وصول هذه الإنزيمات إلى الركيزة [3].
توضح الأدبيات الحديثة حول إنزيمات نزع التفرّع أن pullulanase يرتبط صناعيًا بتعديل النشا وإنتاج شرابات السكر لأنه يستهدف رابطة تختلف عن الرابطة التي تهيمن عليها إنزيمات الأميليز التقليدية. هذه الخصوصية مهمة تجاريًا: فالهدف في مصنع الشراب ليس مجرد “تحلل النشا”، بل الوصول إلى ملف سكري محدد، مثل شراب غني بالجلوكوز أو شراب غني بالمالتوز، مع أقل قدر ممكن من الدكسترينات الثقيلة التي تؤثر في اللزوجة، الترشيح، والثبات بين الدُفعات [1].
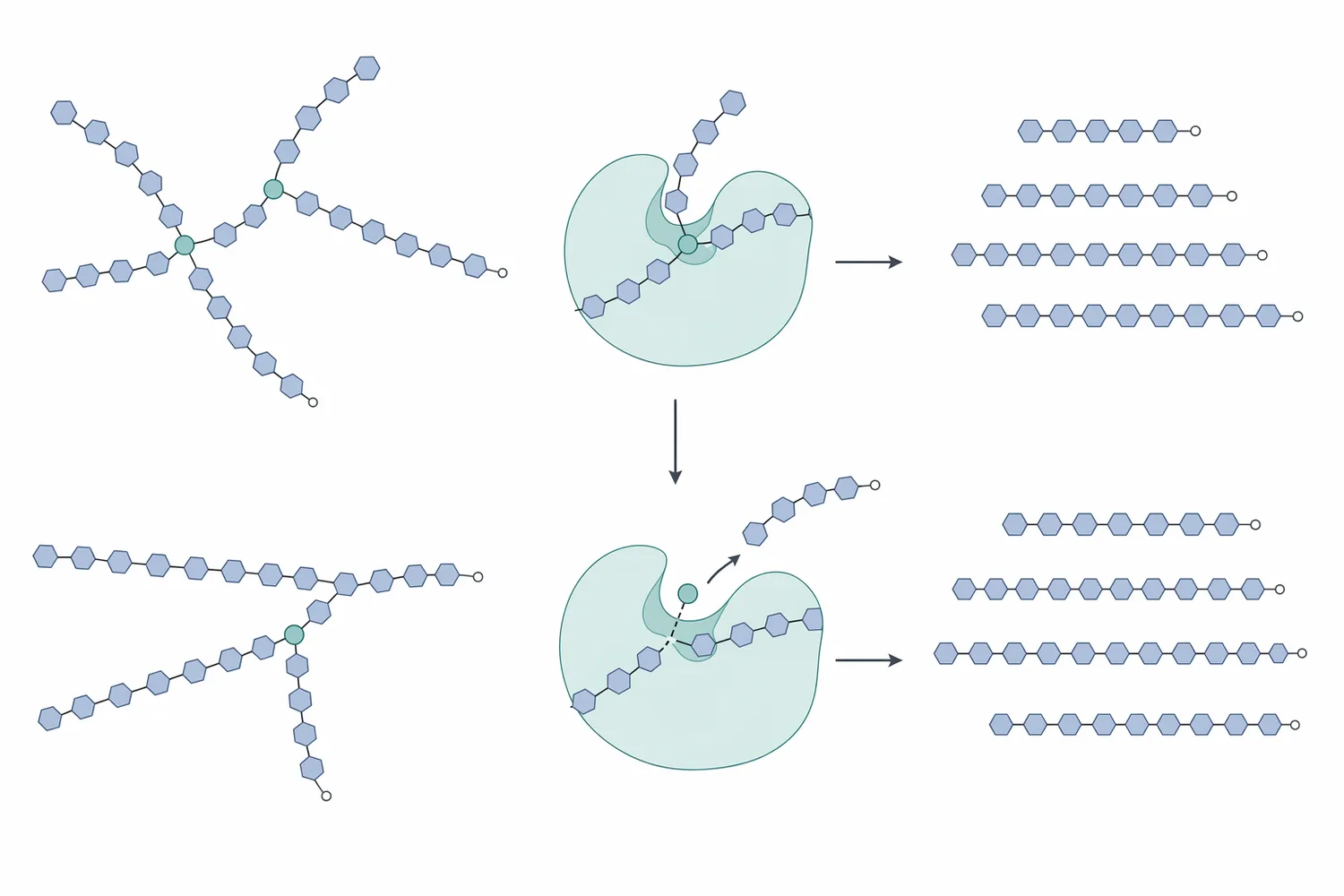
يمكن تشبيه الأميلوبكتين بشجرة كثيفة. الروابط α-1,4 تمثل الامتداد الطولي للفروع، أما روابط α-1,6 فهي نقاط خروج الفروع الجانبية. إنزيمات مثل α-amylase أو glucoamylase أو β-amylase تعمل بكفاءة أكبر على أجزاء معينة من السلاسل، لكنها لا تتعامل مع جميع نقاط التفرّع بالفاعلية نفسها. عندما يُضاف pullulanase إلى وسط نشوي مُسال، فإنه يهاجم نقاط التفرّع، فيحوّل جزءًا من البنية المتشعبة إلى سلاسل أكثر خطية وقابلية لاستكمال السَكَرة [2].
هذا الفعل لا يعني أن pullulanase يزيد “كمية السكر” بطريقة مستقلة عن بقية الإنزيمات، بل يعني أنه يغيّر هندسة الركيزة. بعد إزالة التفرعات، تصبح نهايات السلاسل أكثر إتاحة للجلوكوأميليز في مسار الجلوكوز، أو للإنزيمات المالتيّة في مسار المالتوز. لذلك يكون أثره الأوضح عادةً في انخفاض الدكسترينات المتفرعة وارتفاع قابلية النظام للوصول إلى التركيب السكري المستهدف، لا في إنشاء مسار كيميائي منفصل عن السَكَرة [4].
وتُظهر دراسات خصائص pullulanase أن الإنزيمات من هذا النوع تُقيَّم صناعيًا على أساس الاستقرار، التوافق مع الوسط، وتحمل ظروف المعالجة، لأن التطبيق الفعلي يحدث داخل خليط معقد: دكسترينات بدرجات بلمرة مختلفة، مواد صلبة ذائبة، إنزيمات أخرى، وأيونات أو مكونات قادمة من مصدر النشا. لذلك يجب أن تُفهم الآلية على مستويين: رابطة جزيئية محددة هي α-1,6، ونظام عملية كامل يحدد مدى ترجمة هذا القطع إلى زيادة فعلية في الجلوكوز أو المالتوز [5].
في إنتاج شراب الجلوكوز أو المالتوز، تأتي مرحلة نزع التفرّع عادةً بعد أن يكون النشا قد أصبح في صورة قابلة للمعالجة المائية، أي بعد تقليل البنية الحبيبية واللزوجة وتحويل الكتلة النشوية إلى دكسترينات. عند هذه النقطة لا يكون المطلوب من pullulanase تفكيك حبيبات النشا الأصلية، بل التعامل مع دكسترينات متفرعة نشأت من الأميلوبكتين. هذا التوقيت مهم لأن وصول الإنزيم إلى روابط α-1,6 يكون أفضل عندما تكون الركيزة ذائبة أو مشتتة على نحو يسمح بالتلامس الفعلي بين الإنزيم والرابطة المستهدفة [6].
في مسار شراب الجلوكوز، يُستخدم pullulanase غالبًا بجوار glucoamylase. الجلوكوأميليز يحرر وحدات الجلوكوز من أطراف السلاسل، لكن التفرعات تقلل عدد المسارات المتاحة أو تترك بقايا دكسترينية. بإزالة نقاط α-1,6، يزداد عدد السلاسل الخطية التي يستطيع الجلوكوأميليز متابعتها حتى وحدات أصغر، فتتحسن احتمالية رفع نسبة الجلوكوز في الشراب عند ضبط بقية شروط العملية [4].
أما في مسار شراب المالتوز، فالفكرة مختلفة قليلًا. الهدف ليس الوصول إلى الجلوكوز الحر بأقصى قدر، بل تحرير وحدات مالتوز والمحافظة على ملف سكري مناسب. لذلك يعمل pullulanase كأداة هندسية للركيزة: يزيل التفرعات التي تعيق β-amylase أو الأنظمة المالتيّة، ويتيح سلاسل يمكن أن تُنتج مالتوزًا بدل أن تبقى كدكسترينات متفرعة. ومن هنا تأتي أهمية توافق نظام الإنزيمات؛ فاختيار إنزيمات تدفع بقوة نحو الجلوكوز قد لا يخدم شراب مالتوز مرتفعًا، حتى لو كان نزع التفرّع ناجحًا [1].
| الإنزيم | الهدف البنيوي في النشا | الدور العملي في خط الشراب | الأثر المتوقع على المنتج | هل يحل محل Pullulanase؟ |
|---|---|---|---|---|
| α-amylase | روابط α-1,4 داخلية في السلاسل | تقليل اللزوجة وتكوين دكسترينات قابلة للسَكَرة | يهيئ النشا للمعالجة اللاحقة | لا؛ لأنه لا يزيل عائق α-1,6 بكفاءة كافية في الدكسترينات المتفرعة [3] |
| Glucoamylase | أطراف السلاسل لتحرير الجلوكوز | إنتاج شراب جلوكوز أو دكستروز من الدكسترينات | زيادة الجلوكوز الحر عند توفر سلاسل قابلة للوصول | لا؛ يستفيد من نزع التفرّع لكنه لا يؤدي وظيفة pullulanase نفسها [4] |
| β-amylase والإنزيمات المالتيّة | تحرير وحدات مالتوز من السلاسل المناسبة | إنتاج شراب مالتوز | رفع جزء المالتوز عند ملاءمة الركيزة | لا؛ تحتاج إلى سلاسل أقل تفرعًا لتعمل بكفاءة أعلى [1] |
| Pullulanase | روابط α-1,6 عند نقاط التفرّع | نزع تفرّع الأميلوبكتين والدكسترينات | تقليل الدكسترينات المتفرعة ودعم الجلوكوز أو المالتوز حسب النظام | هو إنزيم داعم متخصص، لا بديلًا كاملًا عن إنزيمات السَكَرة [2] |
هذه المقارنة توضح أن إدخال pullulanase لا يعني زيادة عدد الإنزيمات عشوائيًا، بل معالجة عائق محدد في بنية النشا. فإذا كان التسييل جيدًا لكن الشراب النهائي يحتوي على جزء مرتفع من الدكسترينات المتبقية، فقد يكون السبب بنيويًا مرتبطًا بالتفرعات وليس فقط بكمية الإنزيمات التي تكسر روابط α-1,4. وهنا يصبح نزع التفرّع وسيلة لتحسين قابلية الوصول، وليس مجرد خطوة إضافية في التسلسل [7].
شراب الجلوكوز ينتج من تحويل النشا إلى سكريات مختزلة يكون الجلوكوز فيها مكونًا رئيسيًا بدرجات مختلفة حسب مواصفة الشراب. في الأدبيات، دُرست مصادر نشا متعددة لإنتاج شراب الجلوكوز، مثل القمح، السورغم، البطاطا الحلوة، الكسافا، ومصادر غير تقليدية أخرى. ورغم اختلاف المواد الخام، يتكرر المبدأ نفسه: نجاح التحلل الإنزيمي يعتمد على قابلية النشا للتسييل والسَكَرة، وعلى قدرة الإنزيمات على الوصول إلى الروابط داخل الدكسترينات [8].
في هذا التطبيق، تكون وظيفة pullulanase مباشرة: تقليل الدكسترينات الحدّية المتفرعة التي تحد من اكتمال عمل glucoamylase. عندما يفتح pullulanase نقاط التفرّع، يزداد عدد السلاسل التي يمكن للجلوكوأميليز أن يتعامل معها من الأطراف، ما يدعم رفع الجلوكوز وتقليل السكريات الأعلى غير المرغوبة. لذلك يُستخدم pullulanase كجزء من استراتيجية لتحسين التحويل، وليس كعامل منفرد؛ فالشراب النهائي يتحدد بتوازن نشاط الجلوكوأميليز، قابلية الدكسترينات، زمن المعالجة، وتركيب المادة الصلبة [4].
وتوضح دراسات إنتاج شراب الجلوكوز من نشويات مختلفة أن تركيز الركيزة والإنزيمات، وطبيعة المصدر النشوي، كلها تؤثر في مردود التحلل وفي خصائص الشراب. مثلًا، تناولت أبحاث على نشا السورغم الأحمر ونشا القمح أثر متغيرات العملية على إنتاج شراب الجلوكوز، وهو ما يدعم فكرة أن pullulanase يجب أن يُنظر إليه ضمن منظومة كاملة لا كإضافة تعطي النتيجة نفسها في كل ركيزة [9].
تزيد أهمية نزع التفرّع عندما تكون المادة الخام أكثر تعقيدًا أو عندما تحتوي على بنى مقاومة للتحلل. فالدراسات المتعلقة بمقاومة النشا للتحلل الإنزيمي تبيّن أن البنية الداخلية، الترتيب الجزيئي، وتقييد وصول الإنزيمات يمكن أن تقلل سرعة التحويل حتى لو كانت الروابط الكيميائية نفسها قابلة للتحلل نظريًا. لذلك فإن أي خطوة تزيد إتاحة الروابط، مثل التسييل الجيد ونزع التفرّع، يمكن أن تكون حاسمة في الوصول إلى سَكَرة أعمق [10].
في شراب المالتوز، لا يكون الهدف هو دفع كل السلاسل نحو الجلوكوز، بل تكوين ملف غني بالمالتوز مع ضبط السكريات المصاحبة. هنا يصبح pullulanase مفيدًا لأنه يحرر السلاسل من نقاط التفرّع التي تعيق عمل الإنزيمات المالتيّة. عندما يبقى الأميلوبكتين أو الدكسترينات المتفرعة دون فتح، فإن β-amylase أو الإنزيمات المالتيّة قد تنتج مالتوزًا من الأجزاء المتاحة لكنها تترك دكسترينات متفرعة تحد من نقاء الشراب وترفع جزء السكريات الأعلى [1].
يختلف هذا التطبيق عن شراب الجلوكوز في أن المبالغة في إنزيمات تطلق الجلوكوز قد تغير هدف المنتج. لذلك يكون pullulanase في نظام المالتوز أشبه بمُحسّن لبنية الركيزة لا بمحرك نهائي للتركيب السكري. الإنزيم المالتي هو الذي يحدد اتجاه إنتاج المالتوز، بينما يوفر pullulanase سلاسل أقل تفرعًا وأكثر ملاءمة. هذه النقطة مهمة عند قراءة بيانات الأداء: ارتفاع المالتوز لا ينتج من pullulanase وحده، بل من توافقه مع الإنزيم المالتي وبنية الدكسترينات الناتجة من التسييل [2].
ومن منظور جودة الشراب، يؤدي تقليل الدكسترينات المتفرعة إلى تحسين الاتساق العملي. الدكسترينات الأعلى قد تؤثر في اللزوجة، الترشيح، وسلوك التركيز، كما أنها تغير ملف السكريات النهائي. لذلك فإن نزع التفرّع في شراب المالتوز يخدم هدفين معًا: رفع قابلية تكوين المالتوز وتقليل المكونات المتبقية التي لا تتوافق مع مواصفات الشراب المطلوبة [7].
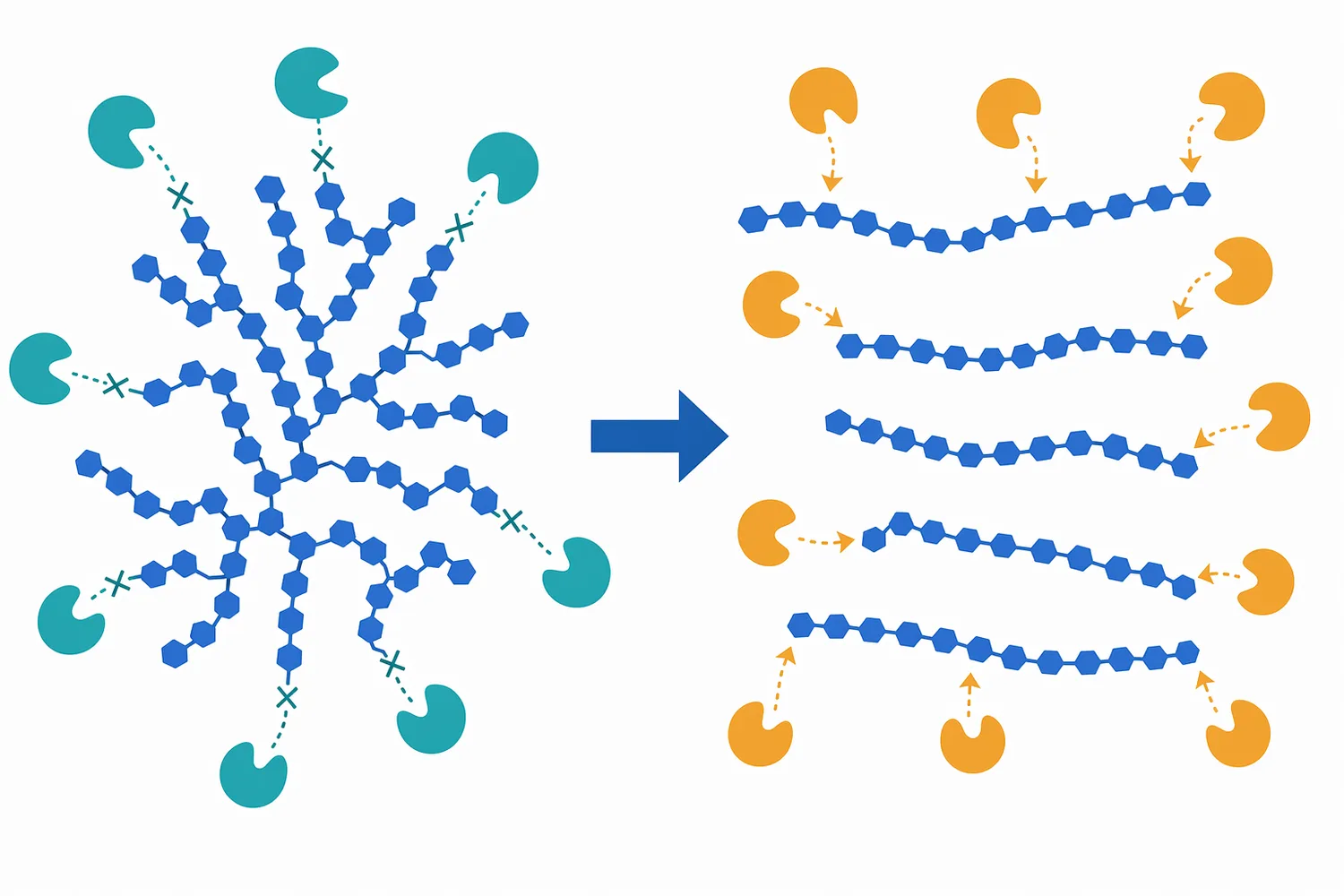
ليس كل نشا يتصرف بالطريقة نفسها. نشا الذرة أو القمح أو الكسافا أو البطاطا الحلوة أو المصادر غير التقليدية يختلف في نسبة الأميلوز إلى الأميلوبكتين، حجم الحبيبات، وجود مكونات مصاحبة، وسهولة التحول إلى دكسترينات قابلة للسَكَرة. لذلك فإن دور pullulanase قد يكون أكثر وضوحًا في أنظمة يغلب عليها تأثير الأميلوبكتين أو تظهر فيها دكسترينات متفرعة بعد التسييل [8].
المراجعات المنهجية حول إنتاج شراب الجلوكوز بالطرق الإنزيمية والحمضية تشير إلى أن الطرق الإنزيمية تقدم انتقائية أعلى في توجيه الناتج، بينما يعتمد الأداء على مصدر النشا وتصميم المعالجة. في هذا السياق، لا يُستخدم pullulanase لتجاوز الحاجة إلى تسييل مناسب أو ضبط العملية، بل لإضافة خطوة انتقائية تستهدف رابطة لا تتعامل معها إنزيمات السَكَرة الرئيسية بالفاعلية نفسها [3].
كما أن دراسات على نشويات مثل البطاطا الحلوة والقلقاس والكسافا وغيرها تؤكد أن تحسين إنتاج الشراب غالبًا يتطلب فهم تفاعل عوامل متعددة، لا مجرد اختيار إنزيم واحد. نوع الركيزة، مستوى التفكك السابق، وتركيب الدكسترينات كلها تحدد ما إذا كان نزع التفرّع سيظهر أثره بوضوح في ارتفاع السكر المستهدف أو انخفاض الدكسترينات [11].
فاعلية pullulanase تتأثر بمدى جاهزية الركيزة. فإذا كان التسييل غير كافٍ وبقيت بنية النشا كثيفة أو غير متاحة، فلن يكون وصول الإنزيم إلى روابط α-1,6 مثاليًا. وإذا كان التسييل شديدًا بطريقة تنتج ملف دكسترينات غير مناسب للهدف، فقد لا يترجم نزع التفرّع إلى الزيادة المتوقعة في الجلوكوز أو المالتوز. لذلك يعتمد الأداء على التسلسل الكامل للعملية: تحضير النشا، التسييل، السَكَرة، ثم المعالجة اللاحقة للشراب [6].
التوافق بين الإنزيمات عامل حاسم أيضًا. في شراب الجلوكوز، يجب أن يعمل pullulanase في بيئة لا تعيق glucoamylase، لأن الفائدة تأتي من إزالة التفرعات ثم تحويل السلاسل الناتجة إلى جلوكوز. في شراب المالتوز، يجب أن يكون النظام موجهًا إلى المالتوز لا إلى تكسير زائد للمالتوز نفسه أو تحويل غير مرغوب نحو الجلوكوز. هذا يفسر لماذا تُصمم أنظمة الشراب عادةً كمزيج إنزيمي وظيفي، لا كمجموعة إنزيمات مستقلة [4].
كما تلعب الشوائب والمكونات غير السكرية دورًا في جودة الشراب النهائي. فقد ناقشت الأدبيات الحديثة حول هيدروليزات النشا أن الشرابات الصناعية لا تحتوي فقط على سكريات مستهدفة، بل قد تضم أملاحًا، بقايا عالية الوزن الجزيئي، أو مكونات تؤثر في التنقية والاستقرار. تقليل الدكسترينات المتفرعة بإنزيم نزع التفرّع لا يغني عن خطوات المعالجة اللاحقة، لكنه يمكن أن يجعل ملف الهيدروليزات أقرب إلى الهدف قبل التنقية أو التركيز [7].
الفائدة الأولى هي تحسين قابلية السَكَرة. عندما تُزال روابط α-1,6، تتحول البنية المتفرعة إلى سلاسل يمكن للإنزيمات الأخرى التعامل معها بشكل أفضل. هذا ينعكس في شراب الجلوكوز كدعم لعمل glucoamylase، وفي شراب المالتوز كدعم للإنزيمات المالتيّة. لذلك يرتبط pullulanase بتحسين التحويل لا من خلال إنتاج السكر النهائي وحده، بل من خلال زيادة إتاحة الركيزة [2].
الفائدة الثانية هي خفض الدكسترينات المتبقية. الدكسترينات المتفرعة تمثل جزءًا من الكربوهيدرات التي لم تصل إلى السكر المستهدف، وقد تؤثر في اللزوجة والخواص الفيزيائية للشراب. بإزالة التفرعات، يمكن تقليل هذا الجزء عند توافق العملية، ما يدعم جودة أكثر اتساقًا بين الدُفعات ويقلل الحاجة إلى تعويض مشكلات البنية المتفرعة في المراحل اللاحقة [7].
الفائدة الثالثة هي المرونة بين شراب الجلوكوز وشراب المالتوز. نفس مبدأ نزع التفرّع يخدم هدفين مختلفين بحسب الإنزيم المصاحب: مع glucoamylase يدعم الجلوكوز، ومع إنزيم مالتي يدعم المالتوز. هذه المرونة لا تعني أن المنتج يعطي كلا الهدفين في العملية نفسها دون تصميم مناسب، بل تعني أن pullulanase أداة مشتركة في مسارات سَكَرة مختلفة [1].
الفائدة الرابعة هي دعم استخدام أوسع لمصادر النشا. مع تزايد الاهتمام بالنشويات غير التقليدية ومخلفات المعالجة الغنية بالنشا، تصبح الإنزيمات الانتقائية مهمة لتحويل مواد خام متنوعة إلى شرابات قابلة للاستخدام. المراجعات الحديثة حول تحويل المواد النشوية غير التقليدية تؤكد أن الاختلاف في بنية الركيزة يستدعي حلولًا إنزيمية أكثر دقة، ومن بينها إنزيمات نزع التفرّع [8].
Pullulanase لا يعوض ضعف التسييل الأولي. إذا بقيت كتلة النشا عالية اللزوجة أو غير متاحة، فإن إنزيم نزع التفرّع لن يؤدي وحده إلى شراب عالي التحويل. وظيفته تبدأ عندما تكون الدكسترينات المتفرعة في صورة يستطيع الإنزيم الوصول إليها. لذلك، في التطبيقات الصناعية، يجب أن يكون جزءًا من سلسلة معالجة متوازنة تبدأ من إعداد المادة الخام وتنتهي بتنقية الشراب أو تركيزه حسب المنتج المطلوب [3].
كما أنه لا يحدد وحده ما إذا كان المنتج النهائي شراب جلوكوز أو شراب مالتوز. هذا القرار ينتج من الإنزيم الرئيسي المصاحب ومن تصميم السَكَرة. إذا كان النظام موجهًا بالجلوكوأميليز، فإن فتح التفرعات سيدعم تحرير الجلوكوز. وإذا كان النظام موجهًا بإنزيمات مالتيّة، فإن فتح التفرعات سيدعم تكوين المالتوز. أما استخدام pullulanase دون توافق إنزيمي، فقد يقلل بعض التفرعات لكنه لا يضمن ملفًا سكريًا مطابقًا للهدف [4].
ولا ينبغي تفسير الأدلة المنشورة على أنها ضمان لقيمة رقمية موحدة في كل عملية. الدراسات على إنتاج شرابات النشا تُظهر باستمرار أن الأداء يتغير حسب الركيزة، تركيبة الإنزيمات، حالة الدكسترينات، وزمن المعالجة. لذلك فالحديث الفني الأدق هو أن pullulanase يعالج عائقًا معروفًا في البنية النشوية، وأن حجم التحسن يعتمد على مدى كون ذلك العائق هو العامل المحدد في النظام المعين [11].
بصفته إنزيمًا سائلًا، يكون Pullulanase مناسبًا للدمج في أنظمة مائية لمعالجة النشا، حيث يساعد الشكل السائل على التوزيع في الوسط عند وجود خلط مناسب. لكن الإنزيمات عمومًا بروتينات فعالة حيويًا، ويجب التعامل معها مهنيًا وفق وثائق السلامة المصاحبة، خاصة عند المناولة، منع الرذاذ، وتجنب التعرض غير الضروري. لذلك تُعد وثيقة SDS جزءًا عمليًا من الاستخدام المسؤول، وليست مجرد مستند إداري .
أما CoA فيدعم تتبع الدفعة ومطابقة بيانات المنتج المرفقة بالطلب. وبما أن Enzymes.bio جهة توريد وليست جهة تصنيع أو مختبرًا، فإن دورها هو إتاحة المنتج ووثائقه المصاحبة للشراء المباشر عبر الإنترنت بوحدة 1 كجم. لا ينبغي قراءة هذه الوثائق على أنها تصميم عملية جاهز، بل كجزء من حزمة معلومات المنتج التي يستخدمها العميل ضمن نظامه التشغيلي الداخلي .
إنزيم Pullulanase Enzyme Liquid For Starch Hydrolysis In Glucose And Maltose Syrup Production هو أداة نزع تفرّع موجهة إلى روابط α-1,6 في الدكسترينات المتفرعة الناتجة من النشا. أهميته العملية أنه يفتح البنية أمام إنزيمات السَكَرة: يدعم glucoamylase في شراب الجلوكوز، ويدعم الإنزيمات المالتيّة في شراب المالتوز، مع أثر متوقع يتمثل في تقليل الدكسترينات المتبقية وتحسين الوصول إلى ملف سكري أكثر اتساقًا [1].
تكون أفضل قراءة لهذا المنتج أنه مكوّن داخل منظومة تحلل النشا، لا بديلًا عن التسييل أو عن إنزيمات إنتاج الجلوكوز والمالتوز. تختلف النتيجة بحسب مصدر النشا، بنية الدكسترينات، وتوافق الإنزيمات المصاحبة، لكن الأساس الميكانيكي ثابت: إزالة التفرعات تزيد قابلية السلاسل للتحويل. المنتج متاح من Enzymes.bio للشراء المباشر عبر الإنترنت بوحدة 1 كجم، مع إرفاق CoA وSDS مع الطلب .
يُباع بوحدة 1 kg، وهو متوفر في المخزون وجاهز للشحن. اطلب مباشرة من متجرنا — ادفع عبر الإنترنت وسنعالج طلبك. تُرفق شهادة التحليل ونشرة بيانات السلامة مع كل طلب.
اشترِ Pullulanase Enzyme Liquid For Starch Hydrolysis In Glucose And Maltose Syrup Production →مرقّمة حسب ترتيب أول اقتباس. مصادر مفتوحة الوصول، تم التحقق من إتاحتها عند النشر؛ وترتبط أرقام الاستشهاد في النص هنا.