Ribonuclease, often abbreviated RNase, is an enzyme used to degrade RNA by cutting the phosphodiester backbone that holds ribonucleotides together. In practical workflows, it is most useful when RNA is an unwanted impurity—such as in DNA extraction, plasmid preparation, genomic DNA preparation, lysate cleanup, or recombinant protein processing—because long RNA molecules can affect viscosity, purity, quantification, and downstream handling. Enzymes.bio supplies Ribonuclease directly online in 1 kg units, with a Certificate of Analysis and Safety Data Sheet included with the order.
Ribonuclease is not one single biological molecule but a broad enzyme class whose members all act on RNA. A ribonuclease protein recognizes RNA as a substrate and catalyzes cleavage of chemical bonds in the RNA backbone, converting long RNA strands into shorter oligonucleotides or nucleotide-ended fragments. This is why the simplest answer to “what does ribonuclease do?” is: it breaks down RNA into smaller pieces by enzymatic cleavage rather than by nonspecific chemical destruction [1].
For process use, the value is straightforward: RNA often behaves as an impurity when the target material is DNA, plasmid DNA, a clarified lysate, or a recombinant protein. Long RNA chains can increase solution viscosity, carry through purification steps, bind proteins or nucleic-acid matrices, and complicate interpretation of nucleic-acid measurements. Ribonuclease is therefore used where intentional RNA degradation improves the manageability or analytical cleanliness of a material stream [2].
The same property also creates an important boundary. In workflows where intact RNA is the product or analytical target—such as RNA therapeutics, RNA delivery studies, in vitro transcription outputs, or RNA-preservation work—ribonuclease is not a helpful additive but a contamination risk. RNase-control guidance emphasizes that RNases are common, persistent contaminants in RNA work, which is why RNA-focused environments treat them as something to exclude rather than introduce [3].
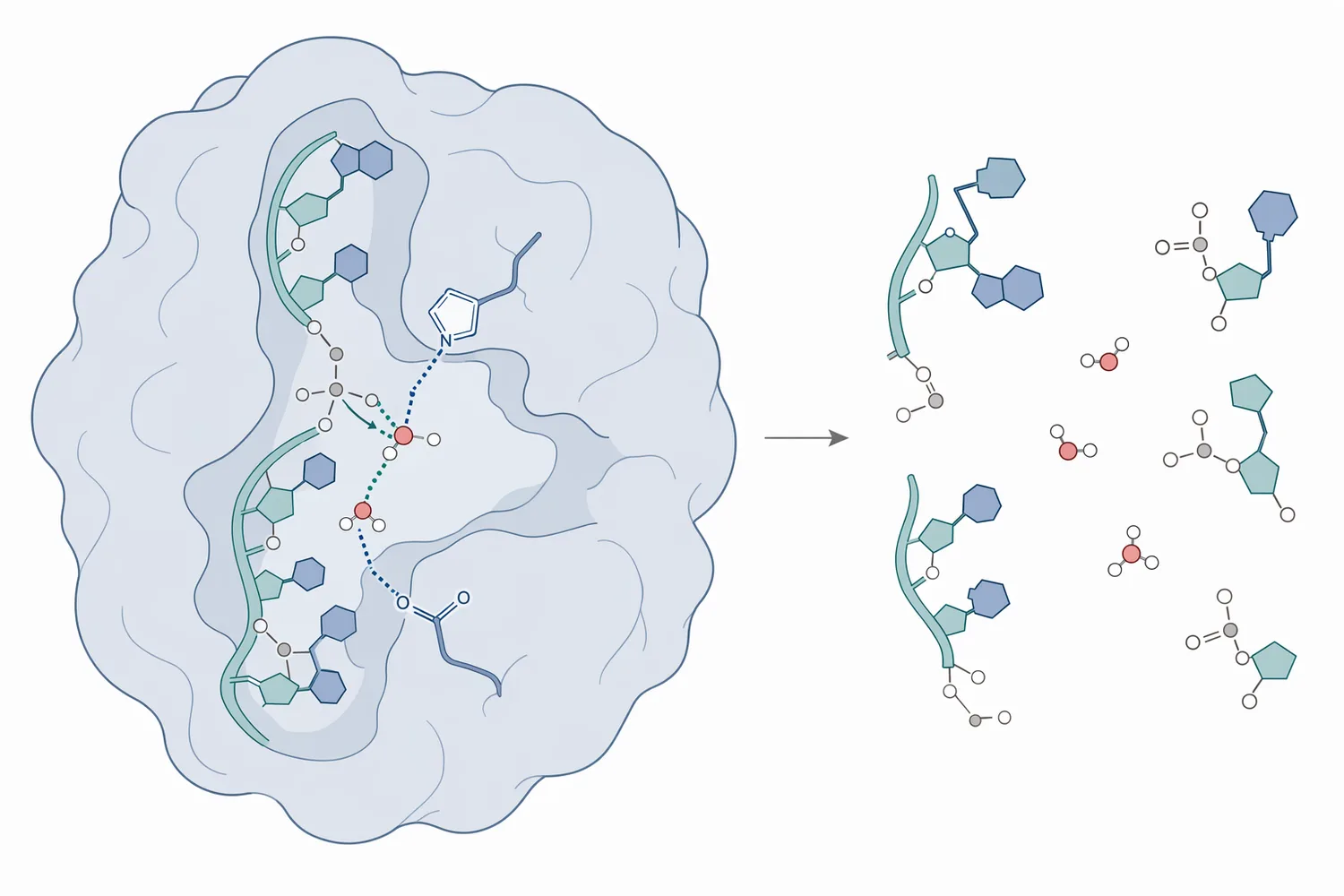
RNA is a polymer made from ribonucleotides joined by phosphodiester bonds. Each linkage connects the sugar-phosphate backbone of one nucleotide to the next, creating a long, flexible, negatively charged chain. Ribonuclease works by binding RNA and catalyzing cleavage of these backbone linkages, so the physical result is shorter RNA fragments with new phosphate-containing ends rather than intact, high-molecular-weight RNA [1].
The best-known example is ribonuclease A, often written RNase A. The ribonuclease A mechanism is widely used as a model for explaining RNase chemistry because the enzyme has been studied for decades as a compact, stable, single-chain protein. RNase A preferentially cleaves single-stranded RNA on the 3′ side of pyrimidine residues, and the reaction proceeds through a cyclic phosphate intermediate before hydrolysis produces phosphate-ended RNA fragments [2].
Mechanistically, the important feature is that RNA contains a 2′-hydroxyl group on the ribose sugar, while DNA does not. RNase A uses that 2′-hydroxyl as an internal nucleophile during cleavage: the RNA strand effectively participates in cutting its own backbone once the enzyme positions the substrate and catalytic residues correctly. This explains why RNase A can target RNA selectively without simply behaving as a general nuclease for all nucleic acids [2].
At the material level, this cleavage changes several practical properties at once. A long RNA molecule can entangle in solution, contribute to viscosity, and remain associated with proteins, membranes, or DNA-containing fractions. After ribonuclease treatment, the same RNA mass is distributed into shorter fragments that are less polymeric and less structurally persistent, which can make downstream clarification, separation, and interpretation easier when RNA removal is the intended operation [1].
Chemical studies on RNase A also show why preserving the active-site environment matters. Work examining chemical modification of ribonuclease A has used the enzyme as a model for how changes to amino-acid side chains can affect protein behavior and biological implications, reinforcing that RNase activity depends on folded protein structure rather than merely the presence of a catalytic chemical group in solution [4].
Technical readers often see names such as ribonuclease A, ribonuclease H, ribonuclease P, ribonuclease T1, ribonuclease III, ribonuclease R, ribonuclease L, and ribonuclease T2. These names are not interchangeable labels for the same enzyme. They describe enzymes or enzyme families with different substrate preferences, biological roles, and cleavage patterns [1].
The distinction between endoribonucleases and exoribonucleases is especially useful. Endoribonucleases cut within an RNA strand, creating internal breaks. Exoribonucleases degrade RNA progressively from an end, removing nucleotides or short fragments from the 5′ or 3′ terminus. For RNA removal from a complex mixture, either type can reduce RNA integrity, but the fragment pattern and substrate requirements may differ substantially [1].
| Ribonuclease type | Typical substrate logic | Main practical meaning |
|---|---|---|
| Ribonuclease A / RNase A | Cleaves single-stranded RNA, especially adjacent to pyrimidines | Common reference enzyme for removing RNA from DNA-oriented and protein-oriented preparations [2] |
| Ribonuclease T1 | Known for guanine-associated RNA cleavage behavior | Useful example of base-preference among RNases |
| Ribonuclease T2 | Broadly distributed RNase family in plants and other organisms | Relevant to biological RNA turnover and plant reproductive systems [5] |
| Ribonuclease H | Acts on RNA in RNA/DNA hybrids | Important conceptually where RNA is paired with DNA rather than free single-stranded RNA |
| Ribonuclease III / ribonuclease 3 | Associated with double-stranded RNA processing | Illustrates that some RNases recognize RNA structure, not just sequence |
| Ribonuclease P | RNA-processing ribonucleoprotein system in many organisms | Important in biology and molecular identification; RNase P RNA genes have been used for bacterial dereplication [6] |
| Ribonuclease R | Exoribonuclease that degrades RNA from an end | Shows how RNA can be removed progressively rather than by internal cuts |
| Ribonuclease L | Immune-linked RNase activated in antiviral pathways | Demonstrates that RNases also function in host defense, not only processing |
This table is conceptual rather than a product-selection matrix. Its purpose is to show why “ribonuclease” is a family term. In commercial RNA-removal work, RNase A is the most familiar name because it is robust, deeply characterized, and commonly associated with removal of RNA from DNA preparations and other non-RNA target materials [2].
DNA extraction often releases RNA at the same time as genomic DNA. If RNA remains in the preparation, it can inflate apparent nucleic-acid recovery, alter absorbance-based estimates, and make the preparation look more abundant or less clean than it really is. Ribonuclease treatment addresses this by degrading RNA while leaving the DNA polymer as the intended nucleic-acid target [2].
The mechanism matters because the goal is not simply “nuclease treatment”; the goal is selective removal of RNA interference. RNase A exploits RNA chemistry, especially the ribose 2′-hydroxyl, to cleave RNA backbones. DNA lacks that same 2′-hydroxyl, which is one reason RNase A is routinely associated with DNA-preparation workflows rather than being treated as a general DNA-degrading enzyme [2].
In genomic DNA workflows, RNA can also affect physical handling. High-molecular-weight RNA contributes to a viscous, stringy matrix after cell lysis, especially in biological samples rich in nucleic acids. Once ribonuclease cuts RNA into shorter fragments, those fragments exert less polymer-like drag in solution, which can support clearer phase behavior, easier mixing, and cleaner downstream separation when the workflow is designed for RNA removal [1].
Plasmid preparation is a classic setting for ribonuclease use because bacterial cells contain abundant RNA. After lysis, plasmid DNA is mixed with chromosomal DNA, RNA, proteins, lipids, salts, and cell-wall components. RNA is not the desired product, but it can remain soluble and carry through intermediate steps unless it is enzymatically shortened [2].
In this context, ribonuclease improves the process by changing RNA from long, interfering molecules into shorter fragments that are easier to separate from plasmid DNA during purification. The enzyme does not “purify plasmid” by itself; instead, it removes one major class of impurity that otherwise complicates the purification train. This is why RNase treatment is typically discussed as a supporting step in plasmid and DNA preparation rather than as a complete purification method [2].
The same logic applies to bacterial or microbial lysates used for other purposes. A study on extracellular production of an RNA aptamer by ribonuclease-free marine bacteria demonstrates the opposite design principle: when RNA is the intended product, organisms and processes may be engineered specifically to avoid ribonuclease degradation. That contrast highlights why RNase is valuable in plasmid DNA cleanup but undesirable in RNA-product manufacture [7].
RNA can be an impurity in recombinant protein preparations, especially when the target protein is expressed in cells and recovered from a nucleic-acid-rich lysate. RNA may remain in the soluble phase, bind to positively charged protein regions, associate with nucleic-acid-binding proteins, or contribute to viscosity that slows clarification and filtration. Ribonuclease reduces this burden by breaking RNA into smaller fragments that are less likely to behave as large co-purifying polymers [1].
For process engineers, the useful mental model is substrate conversion. Before treatment, RNA may be a long polyanion with many possible contact points for proteins, membranes, and purification media. After RNase action, the mixture contains shorter RNA fragments with fewer continuous binding sites and lower chain length. This can reduce RNA’s contribution to non-target interactions, although the final outcome depends on the process matrix and purification design [2].
RNase A is also a prominent model protein in protein-engineering and modification studies. For example, mono-PEGylation work on ribonuclease A examined how chemical modification of the enzyme could alter properties while retaining the identity of RNase A as a catalytic protein model. Such studies are useful because they reinforce that ribonuclease performance is tied to folded protein architecture and accessible catalytic surfaces [8].
In bioprocessing, ribonuclease is most relevant where RNA is an impurity rather than a product. That can include lysate treatment, reduction of nucleic-acid burden in intermediate streams, cleanup of protein-containing fractions, and preparation of DNA-focused materials. The practical benefit comes from cutting RNA’s backbone so it no longer behaves like a long, high-molecular-weight contaminant [2].
Extracellular RNase production has been reported in microorganisms such as Pseudomonas maltophilia, showing that ribonuclease activity is not confined to purified laboratory reagents but is also part of microbial biology and extracellular nutrient processing. This biological ubiquity helps explain why RNase activity is so commonly encountered in biological materials and why RNA-preserving operations must actively guard against it [9].
RNA-based manufacturing illustrates the sharp dividing line between use and avoidance. In one direction, adding ribonuclease can intentionally eliminate RNA from DNA or protein streams. In the other direction, RNase-free systems have been proposed for extracellular production of RNA aptamers, because the desired RNA would otherwise be degraded by the same class of enzyme that is useful in cleanup applications [10].
RNA delivery research also depends on protecting RNA from degradation. Studies on RNA–polymer complexes are built around the reality that RNA must be stabilized and delivered without being prematurely destroyed. That context reinforces the practical rule: ribonuclease belongs in workflows where RNA removal is intentional, not in workflows where RNA integrity is the value of the product [11].
Ribonuclease A deserves special attention because it is one of the most studied proteins in enzymology. It has been used as a reference protein for folding, catalytic mechanism, structural modification, and stability research. Its compact structure and strong historical literature base make it the ribonuclease many technical readers recognize first [8].
The ribonuclease A mechanism is often described in two chemical phases. First, the enzyme promotes transphosphorylation, in which the RNA 2′-hydroxyl attacks the adjacent phosphorus center and creates a 2′,3′-cyclic phosphate intermediate. Second, hydrolysis opens that cyclic phosphate to form a phosphate-ended RNA fragment. The practical result is internal cleavage of RNA strands into shorter fragments [2].
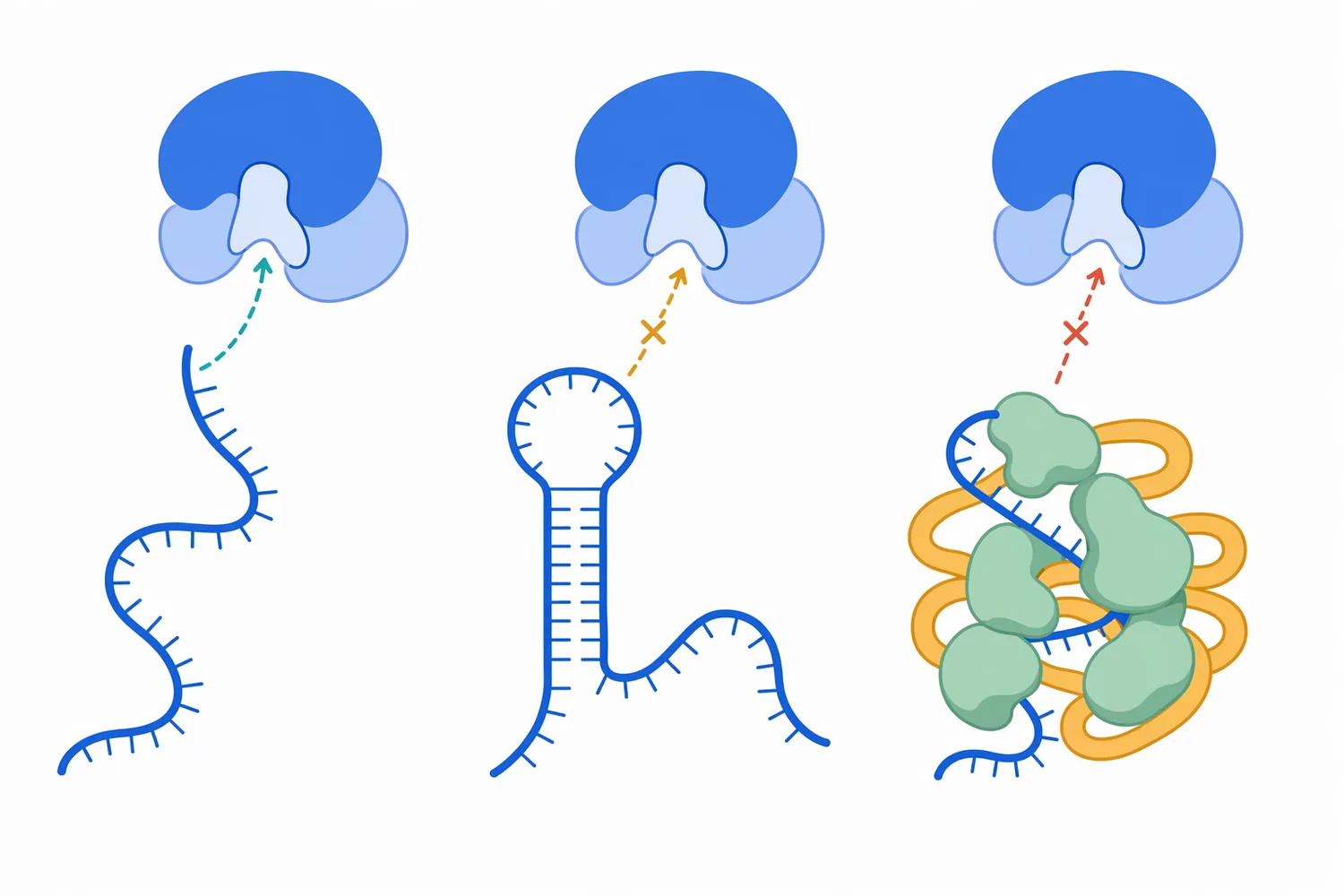
This mechanism also explains why substrate accessibility matters. RNase A acts on RNA regions it can bind and position in the active site. Highly structured RNA, RNA tightly bound by proteins, RNA embedded in complexes, or RNA protected by delivery materials may be less accessible than free single-stranded RNA. In practical terms, the enzyme acts on exposed substrate; it does not magically reach every nucleotide if the matrix physically shields the RNA [1].
Chemical studies reinforce the importance of protein structure and substrate positioning. Work on the biological implications of reactive compounds toward ribonuclease A used RNase A as a target for understanding how chemical interactions with a protein can affect function. For users, the takeaway is not a new handling protocol, but a mechanistic point: RNase activity is a property of a folded enzyme with specific catalytic residues and substrate-binding geometry [4].
While RNase A dominates many practical discussions, other ribonucleases are central to biology. Ribonuclease P is involved in RNA processing and is important enough that RNase P RNA gene sequencing has been evaluated as a tool for molecular dereplication of myxobacterial strain collections. That use reflects the conserved and informative nature of RNase P-related sequences in microbial classification work [6].
Ribonuclease T2 is another family with biological significance. A transcriptome study in the bast fibre crop Crotalaria juncea identified T2 ribonuclease genes to investigate late-acting pollen self-rejection, linking RNase biology to plant reproductive compatibility. This is far removed from DNA cleanup, but it shows how broadly ribonuclease mechanisms appear across living systems [5].
Ribonuclease III, sometimes searched as ribonuclease 3, is generally associated with double-stranded RNA processing rather than the same substrate behavior as RNase A. Ribonuclease H acts on RNA strands within RNA/DNA hybrids, making it conceptually important when RNA is paired with DNA. Ribonuclease R and ribonuclease L illustrate additional biological roles, including exonucleolytic RNA degradation and immune-linked RNA cleavage pathways [1].
These differences matter because the word “ribonuclease” should not be read as a promise that every RNase behaves identically. A nuclease that recognizes double-stranded RNA, a nuclease that acts on RNA/DNA hybrids, and a nuclease that cleaves exposed single-stranded RNA can all belong to the same broad category while performing different jobs. For most RNA-removal applications in DNA and protein workflows, RNase A remains the familiar reference point [2].
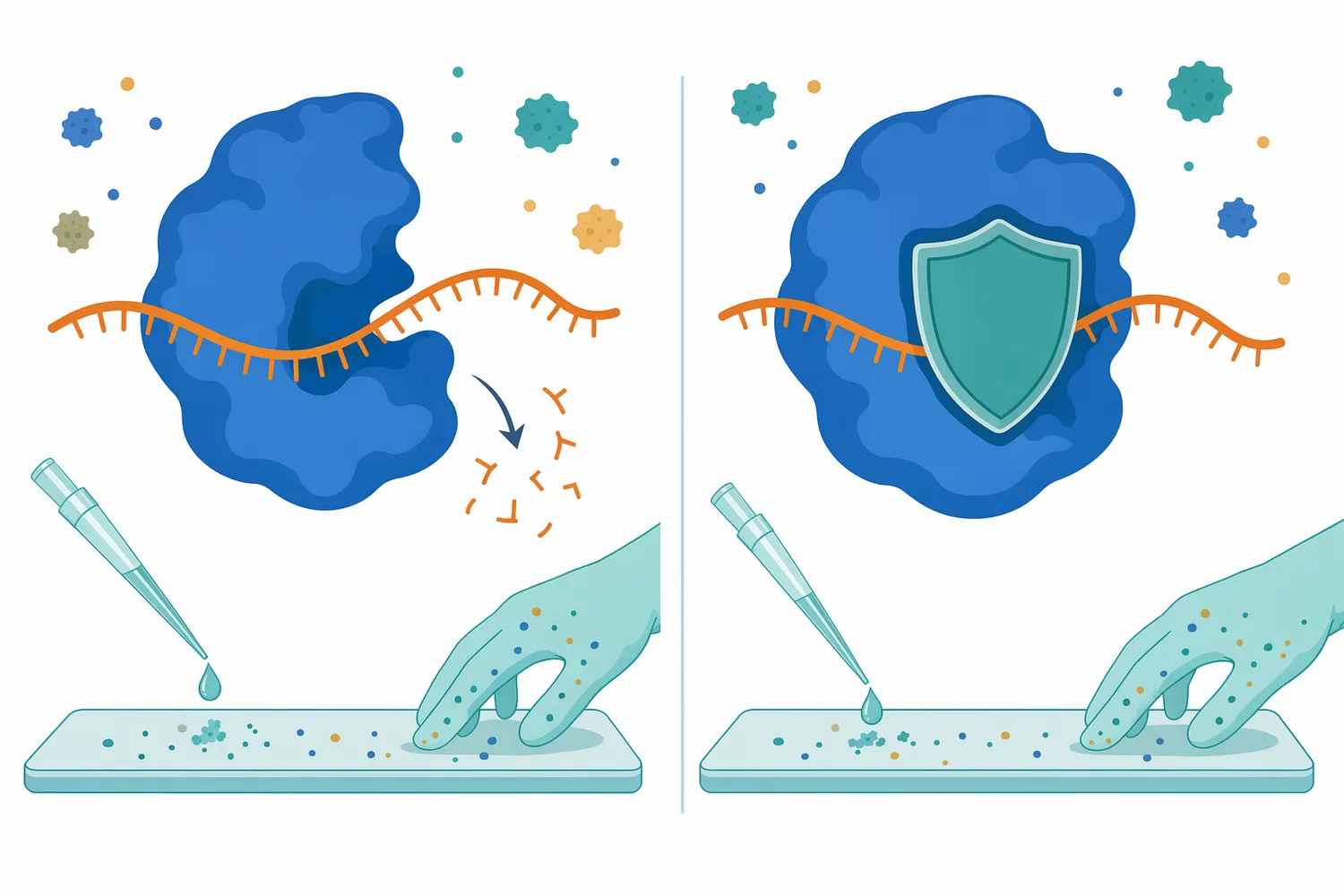
A ribonuclease inhibitor is a molecule that reduces or blocks RNase activity. In biological systems and RNA-preserving workflows, a ribonuclease inhibitor protein may be used to protect RNA from degradation by specific RNases. The concept is the mirror image of intentional RNA removal: instead of adding RNase to eliminate RNA, the workflow suppresses RNase so RNA remains intact [3].
RNase control is emphasized because RNases are common in the environment and can be difficult to eliminate once introduced. They may come from skin contact, biological materials, microorganisms, dust, untreated surfaces, or contaminated consumables. That ubiquity is why RNA-focused workflows use strict RNase-control practices and why ribonuclease products should be kept conceptually and physically separate from RNA-preservation work [3].
The practical distinction is simple. If the desired output is DNA, plasmid DNA, a protein preparation, or a clarified non-RNA stream, ribonuclease may be a helpful processing aid. If the desired output is intact RNA, ribonuclease exposure can reduce yield, damage integrity, or compromise analytical interpretation. The same enzyme class can therefore be either valuable or harmful depending entirely on the process objective [7].
Controlled enzyme switching and inhibition are also active research topics. A study on an enzyme switch using a complementary polymer pair system examined how polymer interactions can regulate enzyme activity, demonstrating the broader principle that RNase function can be modulated by molecular environment and binding partners. This supports the real-world observation that RNase performance depends on the surrounding matrix, not only on the enzyme name [12].
Ribonuclease activity depends on the enzyme, substrate form, and surrounding solution. RNase A is commonly described as active in aqueous systems and is known for cleaving RNA adjacent to pyrimidine residues through the cyclic phosphate pathway. However, observed performance in a real material stream depends on whether the RNA is exposed, folded, protein-bound, hybridized, or trapped in a complex matrix [2].
Substrate chemistry is central. Free single-stranded RNA presents accessible phosphodiester bonds and the 2′-hydroxyl chemistry required for RNase A-type cleavage. RNA/DNA hybrids, double-stranded RNA, structured RNA, and RNA wrapped in proteins or polymer complexes may require different RNase mechanisms or may simply be less accessible. This is one reason that ribonuclease mechanism discussions are more useful than generic claims about “nucleic-acid removal” [1].
The solution environment can also affect folded protein stability and catalytic behavior. Protein studies using RNase A as a model have explored how solvent environments influence structural properties and stability, reinforcing that enzymes are functional proteins whose activity depends on maintaining an appropriate three-dimensional structure. In production or cleanup contexts, the matrix surrounding the enzyme is therefore part of the outcome [13].
Inhibitors, protective molecules, and competing interactions may reduce apparent activity. A ribonuclease inhibitor protein can bind and suppress certain RNases, while nucleic-acid-binding proteins or delivery polymers can shield RNA from enzymatic attack. These effects do not contradict RNase activity; they show that the substrate must be reachable and chemically compatible with the enzyme’s mechanism [3].
The first benefit is improved nucleic-acid purity in DNA-oriented workflows. When RNA is degraded, DNA preparations are less likely to contain intact RNA polymers that interfere with quantification, gel appearance, enzymatic reactions, or downstream interpretation. This is especially relevant in genomic DNA and plasmid DNA work, where the target molecule is DNA rather than total nucleic acid [2].
The second benefit is improved physical handling of lysates and intermediate streams. Long RNA chains can increase viscosity and contribute to difficult mixing, transfer, clarification, or filtration. Ribonuclease reduces chain length, so RNA becomes less able to behave as a large entangled polymer in solution [1].
The third benefit is cleaner protein-oriented processing when RNA is not part of the desired output. RNA can bind proteins, alter charge behavior, and contribute to non-target material in fractions. Enzymatic shortening of RNA can reduce that burden and support a cleaner path through downstream processing, provided RNA degradation is compatible with the product and process objective [2].
The fourth benefit is analytical clarity. If a signal, viscosity issue, or impurity profile is partly RNA-driven, ribonuclease treatment can reduce the RNA contribution and make the remaining material easier to interpret. This is valuable in controlled workflows where the intended answer is “remove RNA and observe the DNA, protein, or non-RNA fraction more clearly” [1].
Ribonuclease should not be treated as a universal cleanup enzyme. It does not replace clarification, chromatography, filtration, precipitation, extraction, or other separation methods. It performs a specific biochemical conversion: RNA backbone cleavage. The downstream process still determines how the resulting fragments, salts, proteins, and other components are separated or retained [2].
It is also not appropriate where intact RNA must be preserved. RNA aptamers, mRNA, guide RNA, viral RNA, transcriptomic samples, and RNA delivery materials can be damaged by RNase exposure. RNase-free production proposals and RNA-control guidance both emphasize the same point from different angles: RNase is powerful because it degrades RNA, so it must be excluded when RNA integrity is the goal [10].
Even when RNA removal is desired, expectations should remain realistic. Highly protected RNA may be less accessible than free RNA, and complex matrices can affect enzyme contact with the substrate. Ribonuclease works by molecular recognition and catalysis, not by physically extracting every RNA-derived fragment from the system. The separation design after RNA cleavage still matters [1].
Finally, RNase contamination control matters for facilities that handle both RNA-removal and RNA-preservation work. The enzyme’s usefulness in one area can become a liability in another. Clear separation of intent—RNA removal versus RNA protection—is the most important operating principle [3].
Enzymes.bio supplies Ribonuclease as a direct online enzyme product sold by the 1 kg unit. Buyers can purchase online, pay online, and the order is processed and shipped. A Certificate of Analysis and Safety Data Sheet are included with the order for documentation and responsible handling.
For customers using ribonuclease in suitable workflows, the core value is dependable RNA degradation where RNA is an unwanted impurity. The strongest-fit applications are DNA extraction, plasmid preparation, genomic DNA preparation, lysate cleanup, recombinant protein processing, and other non-RNA workflows where reducing RNA burden improves purity, handling, or analytical clarity [2].
Ribonuclease is best understood as a precise biochemical tool: it cuts RNA, and that cut changes RNA from long, interfering polymers into shorter fragments. Used with the right process intent, that mechanism supports cleaner DNA- and protein-oriented workflows. Used in the wrong context, especially where RNA is the product, the same mechanism can be harmful—so the correct framing is intentional RNA removal, not general-purpose sample treatment [3].
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Ribonuclease →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.