Ribonuclease(核糖核酸酶,常寫作 RNase)是一類催化 RNA 磷酸二酯鍵水解的酵素,主要應用於 DNA 樣品中的 RNA 去除、核酸製程中間處理、RNA 代謝研究,以及部分宿主防禦與抗菌機制研究。常見類型包括 ribonuclease A、ribonuclease H、ribonuclease P、ribonuclease III 等;它們的底物選擇性、ribonuclease structure、金屬離子需求與 ribonuclease function 並不相同,不能以單一 RNase 概括所有用途。Enzymes.bio 供應的 ribonuclease enzyme 以 1 kg 單位在線上銷售,CoA 與 SDS 會隨訂單一併提供。
搜尋「ribonuclease中文」或「ribonuclease 中文」時,最常見譯名是「核糖核酸酶」。它不是單一酵素,而是能作用於 RNA 的酵素群;其共同點是辨識 RNA 或 RNA 相關結構,並促進 RNA 骨架中磷酸二酯鍵斷裂。對 B2B 使用者而言,最實務的理解方式是:RNase 是用來「控制、移除或研究 RNA」的工具性酵素,而不是泛用型核酸分解劑。不同 RNase 對單股 RNA、雙股 RNA、RNA/DNA hybrid、前驅 tRNA 或特定序列結構的選擇性差異很大,這也是 ribonuclease substrate 選擇會直接影響製程結果的原因 [1]。
在分子生物與生物製程中,RNase 最常被用於去除不需要的 RNA 背景,例如 DNA 萃取後降低 RNA 殘留、降低黏稠度、改善後續純化表現,或在研究中拆解特定 RNA 結構。另一方面,某些 RNase 也是生物體內 RNA 成熟、RNA 品質控制、免疫調節與抗微生物防禦的一部分;例如脊椎動物 RNase A superfamily 不只參與核酸代謝,也被研究其免疫與抗菌相關活性 [2]。
Ribonuclease 的分類常讓使用者混淆,尤其是 ribonuclease a、ribonuclease h、ribonuclease p 與 ribonuclease iii 這幾個名稱在文獻與產品頁中都很常見。它們的命名不是依照「強弱」排序,而是反映來源、結構家族或底物類型。若把所有 RNase 都視為同一種酵素,可能導致錯誤的底物設定、反應條件或下游控制策略 [1]。
| 類型 | 主要底物或作用對象 | 典型功能重點 | 應用理解 |
|---|---|---|---|
| Ribonuclease A / RNase A | 多作用於單股 RNA,偏好特定嘧啶相關切割位點 | 經典胰臟型 RNase;常作為蛋白質摺疊、酵素機制與 RNA 去除模型 | 常見於 DNA 樣品 RNA 去除與生化研究 |
| Ribonuclease H / RNase H | RNA/DNA hybrid 中的 RNA 鏈 | 去除與 DNA 配對的 RNA;與複製、反轉錄與核酸工程相關 | 適合理解 RNA-DNA hybrid 處理,不等同 RNase A |
| Ribonuclease P / RNase P | 前驅 tRNA 等特定 RNA 結構 | RNA 成熟加工,尤其是 tRNA 5′ 端加工 | 常見於 RNA 加工與基因失活概念研究 |
| Ribonuclease III / RNase III | 雙股 RNA 或雙股 RNA 區域 | 金屬依賴型 RNA 切割;與 RNA 加工、調控 RNA 生成相關 | 適合雙股 RNA 結構與 RNA 調控研究 |
| RNase 7 等宿主防禦 RNase | RNA 與微生物表面/膜相關靶點 | 兼具核酸酶活性、陽離子表面與抗菌相關特徵 | 多屬研究導向,不宜直接等同商業抗菌方案 |
RNase A 是最常被當作「ribonuclease enzyme」代表的類型,尤其是 bovine pancreatic ribonuclease(牛胰臟核糖核酸酶)與 pancreatic ribonuclease 相關文獻非常豐富。RNase A 被廣泛用作酵素催化、蛋白質摺疊與結構功能關係的模型蛋白;其小型、穩定、含二硫鍵的特性,使它在 ribonuclease structure and function 研究中具有經典地位 [3]。
Ribonuclease H 則是另一種概念完全不同的 RNase。RNase H 的重點不是任意清除所有 RNA,而是辨識 RNA/DNA hybrid,並水解其中的 RNA 鏈;原核生物 RNase H 研究顯示,其分子多樣性、底物結合區域與催化機制都與 RNase A 類型不同,常涉及金屬離子協同催化 [1]。
Ribonuclease P 是 RNA 生物學中的特殊案例,因為它與前驅 tRNA 加工密切相關,也曾被用於「以內源性 RNase P 促進特定 RNA 切割」的基因失活概念研究。這類用途偏向分子設計與細胞內 RNA 加工研究,不能簡化為一般樣品處理用 RNase [4]。
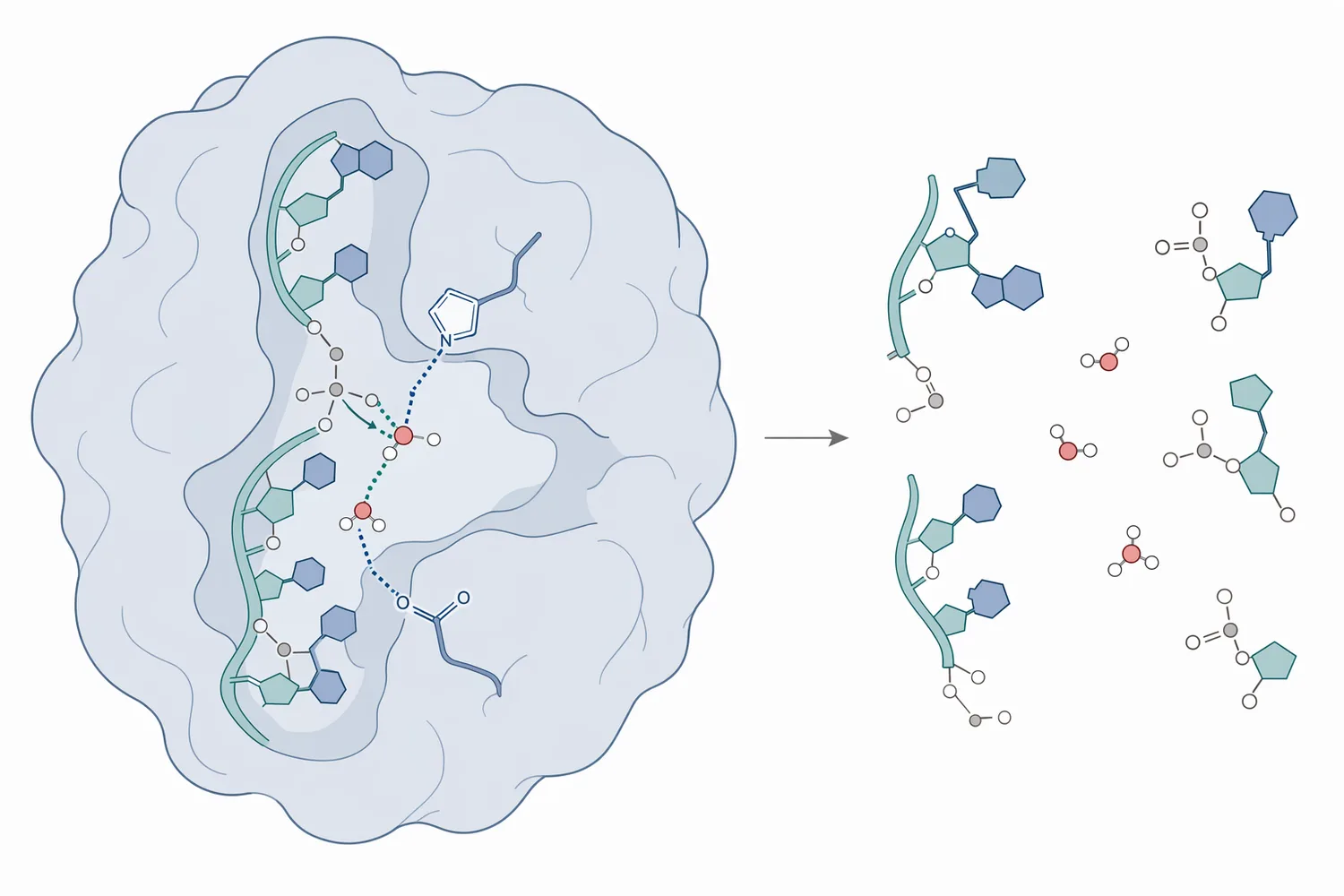
Ribonuclease III 則常與雙股 RNA 結構處理有關。它所代表的是一類可辨識 dsRNA 或局部雙股 RNA 結構的酵素,與 RNA 成熟、RNA 干擾相關通路與調控 RNA 生成有關;在應用理解上,RNase III 更適合被視為「結構性 RNA 處理酵素」,而非一般 RNA 污染去除工具 [1]。
在「ribonuclease a mw」與「ribonuclease a pi」相關搜尋中,使用者多半想確認 RNase A 的基本理化輪廓。商業與文獻資料通常將 RNase A 描述為小型鹼性蛋白,常見來源為 bovine pancreatic ribonuclease;其分子量約在 13.7 kDa 等級,等電點偏鹼性。這些資訊有助於理解它在離子交換、蛋白穩定性與核酸交互作用上的行為,但實際產品文件仍應以隨貨 CoA 與 SDS 為準 [5]。
RNase A 的結構之所以重要,是因為它把「ribonuclease structure」與「ribonuclease function」連結得非常清楚。RNase A 具有緊密摺疊與二硫鍵穩定結構,催化中心位於特定胺基酸殘基形成的凹槽中;底物 RNA 進入後,活性位點會促進磷酸二酯鍵斷裂。RNase A 的蛋白質摺疊研究也常被用來說明變性、復性、二硫鍵重排與天然構形恢復之間的關係 [3]。
RNase A 的催化並非單靠「切割能力」四個字就能描述。點突變研究顯示,位於底物結合與催化環境中的 Lys-7、Arg-10 等殘基會影響酵素的催化特性,尤其是與磷酸結合位點相關的交互作用。這說明 RNase A 對 RNA 的作用取決於活性位點幾何、電荷分布與底物定位,而不只是蛋白質是否能接觸 RNA [6]。
RNase 的基本化學目標,是讓 RNA 中相鄰核苷酸之間的磷酸二酯鍵斷裂。由於 RNA 的核糖具有 2′-羥基,部分 RNase 可利用這個官能基參與轉磷酸化與水解步驟,形成特定末端結構的 RNA 片段。這也是 RNA 比 DNA 更容易在某些條件下發生鹼催化裂解的化學背景,但酵素催化會以更高的選擇性與速率完成同類型反應 [7]。
以 RNase A 類型來看,常見機制包含一般酸鹼催化:活性位點殘基協助活化 2′-羥基,使其攻擊相鄰磷酸,產生環狀磷酸中間體,之後再經水解形成終產物。這個過程仰賴底物在活性中心中的精準定位;若底物結合區被改變,即使酵素整體摺疊仍存在,也可能顯著改變切割效率或反應偏好 [6]。
RNase H 的機制則常以金屬離子協同催化來理解。原核生物 RNase H 研究指出,不同 RNase H 具有多樣的底物結合區與催化架構,但共同關鍵是辨識 RNA/DNA hybrid,並在活性位點中利用金屬離子協助水分子攻擊 RNA 鏈的磷酸骨架。這種底物限制使 RNase H 在分子生物學上具有特別價值:它能處理 hybrid 中的 RNA,而不是任意消化所有核酸 [1]。
RNase 7 與其他宿主防禦 RNase 則提醒我們,RNase 的生物功能不一定只來自 RNA 水解。RNase 7 的結構與調控研究指出,它具有陽離子表面與與微生物互動相關的特性,可能透過與細菌表面或膜結構作用而參與宿主防禦。也就是說,某些 RNase 的抗菌現象可能同時涉及酵素活性、蛋白表面電荷與膜交互作用 [8]。
RNase 活性在某些流程中很有用,但在 RNA 研究、RNA 藥物、mRNA 製程或任何需要保留 RNA 完整性的系統中,RNase 反而是最需要控制的風險之一。Ribonuclease inhibitor 的概念正是來自這種需求:當研究目標是保護 RNA,必須避免外源或內源 RNase 造成降解;當目標是去除 RNA,則要讓 RNase 作用發生在可控步驟,並在後段處理中管理殘留活性 [9]。
酵素抑制不只是「讓酵素失效」,而是透過結合活性位點、改變構形、競爭底物或影響調控區域來降低酵素作用。一般酵素抑制研究顯示,抑制劑可能具有可逆或不可逆特性,也可能因濃度、環境與蛋白構形而改變抑制效果。對 RNase 相關流程而言,抑制策略必須與用途一致:保護 RNA 時重點是避免任何非預期降解;用 RNase 去除 RNA 時重點則是控制作用範圍與後段清除 [9]。
在 DNA 萃取或純化流程中,RNA 殘留常造成樣品黏稠、吸光讀值偏差、下游酶促反應背景增加,或影響濃度估算。使用 RNase A 類型酵素處理 DNA 樣品,是分子生物學中常見的 RNA 去除策略;其價值在於降低 RNA 對 DNA 分析的干擾,而不是提高 DNA 本身的產量。此應用通常需要和後續純化、失活或移除步驟搭配,避免 RNase 在不需要的階段持續存在 [3]。
若樣品中關心的是 RNA/DNA hybrid,則 RNase H 的角色更明確。例如在某些核酸工程、反轉錄相關研究或 hybrid 結構分析中,RNase H 可用來選擇性分解 hybrid 中的 RNA 鏈。這類應用不應與 RNase A 混用概念,因為 RNase H 的底物辨識邏輯是「RNA 與 DNA 配對形成的 hybrid」,而不是一般單股 RNA 污染 [1]。
RNase 不只是處理試劑,也是研究 RNA 生物學的核心工具與研究對象。細胞內 RNA 從轉錄、加工、修飾、定位到降解,每一步都可能涉及 RNase 或 RNase-like 活性。RNase P 在前驅 tRNA 加工中的角色尤其經典,也使其成為 RNA 結構辨識與 RNA 導向切割策略的重要研究模型 [4]。
RNase III 類型則與雙股 RNA 結構處理密切相關,對理解 RNA 調控、非編碼 RNA 成熟與細胞內 RNA 品質控制具有價值。在實務上,若研究目標涉及 dsRNA、髮夾結構或 RNA 調控通路,RNase III 類型的機制會比 RNase A 更接近問題本身。這也是為什麼「ribonuclease function」必須放在具體酵素家族與底物脈絡下討論 [1]。
脊椎動物 RNase A superfamily 的研究顯示,部分 RNase 具有免疫調節與抗微生物相關特性。這些功能不一定能直接轉化為工業抗菌劑,但對研發人員很有啟發性:蛋白質表面正電荷、膜交互作用、RNA 水解活性與局部組織表現,都可能共同影響 RNase 在宿主防禦中的行為 [2]。
RNase 7 是常被討論的例子之一。其結構、調控與宿主防禦貢獻的綜述指出,RNase 7 在皮膚與上皮防禦中具有研究價值,並可能透過對微生物膜或表面結構的作用參與抗菌反應。對 B2B 讀者而言,這類資料適合用於早期研發與機制評估,不應被解讀為現成、已驗證的商業抗菌應用承諾 [8]。
在生物製程或核酸相關材料處理中,RNA 可能是需要控制的雜質、黏度來源或背景干擾。RNase 可在特定中間步驟中協助降低 RNA 含量,尤其是在目標產物不是 RNA、且流程允許後段移除酵素與反應產物的情境中。此處的關鍵不是「加了 RNase 就完成純化」,而是將 RNase 視為整體製程中的一個處理環節,需與分離、純化、殘留控制與品質文件相互配合 [10]。
若考慮固定化或重複使用酵素概念,酵素固定化材料的研究指出,載體性質、孔隙結構、表面官能基與酵素固定方式會影響酵素穩定性、可回收性與操作表現。這類概念可用於理解為何某些製程會評估固定化 RNase 或其他固定化酵素,但實際可行性仍取決於底物、流體條件、清潔需求與最終產品規格 [10]。
RNase 活性受到 pH、溫度、鹽濃度、金屬離子、抑制物與底物結構影響。RNase A 類型通常以相對穩定著稱,但不同來源與配方的穩定性不能直接互推;RNase H 與 RNase III 類型則常涉及金屬依賴性或特定結構底物,因此條件變動對結果的影響可能更明顯。B2B 使用者在導入流程時,應以自身製程條件進行相容性確認,而不是只依賴文獻中的模型條件 [1]。
殘留控制是 RNase 應用中容易被低估的一點。RNase 若殘留在後續需要保護 RNA 的步驟中,可能造成非預期 RNA 降解;若用於 DNA 樣品處理,則需確認後續分析是否會受到蛋白殘留、鹽分或反應副產物影響。這不是 RNase 特有的問題,而是所有酵素製程導入都需要面對的品質控制議題 [7]。
RNase 催化 RNA 水解的能力、RNase A 作為結構與摺疊模型的地位、RNase H 對 RNA/DNA hybrid 的底物選擇性,屬於證據較充分、可重複驗證的基礎知識。這些結論來自長期生化、結構與分子生物研究,適合支撐 DNA 樣品 RNA 去除、RNA 代謝研究與核酸處理流程的技術合理性 [3]。
相較之下,RNase 的抗菌、免疫調節或癌症治療相關應用,雖有活躍研究,但更需要區分「機制研究」與「成熟產品用途」。例如 ribonuclease-based immunotoxins 被討論為抗癌策略之一,其核心概念是利用 RNase 的細胞毒性潛力結合靶向設計,但此類研究涉及遞送、免疫原性、選擇性與安全性等複雜問題,不能直接等同一般工業 RNase 應用 [11]。
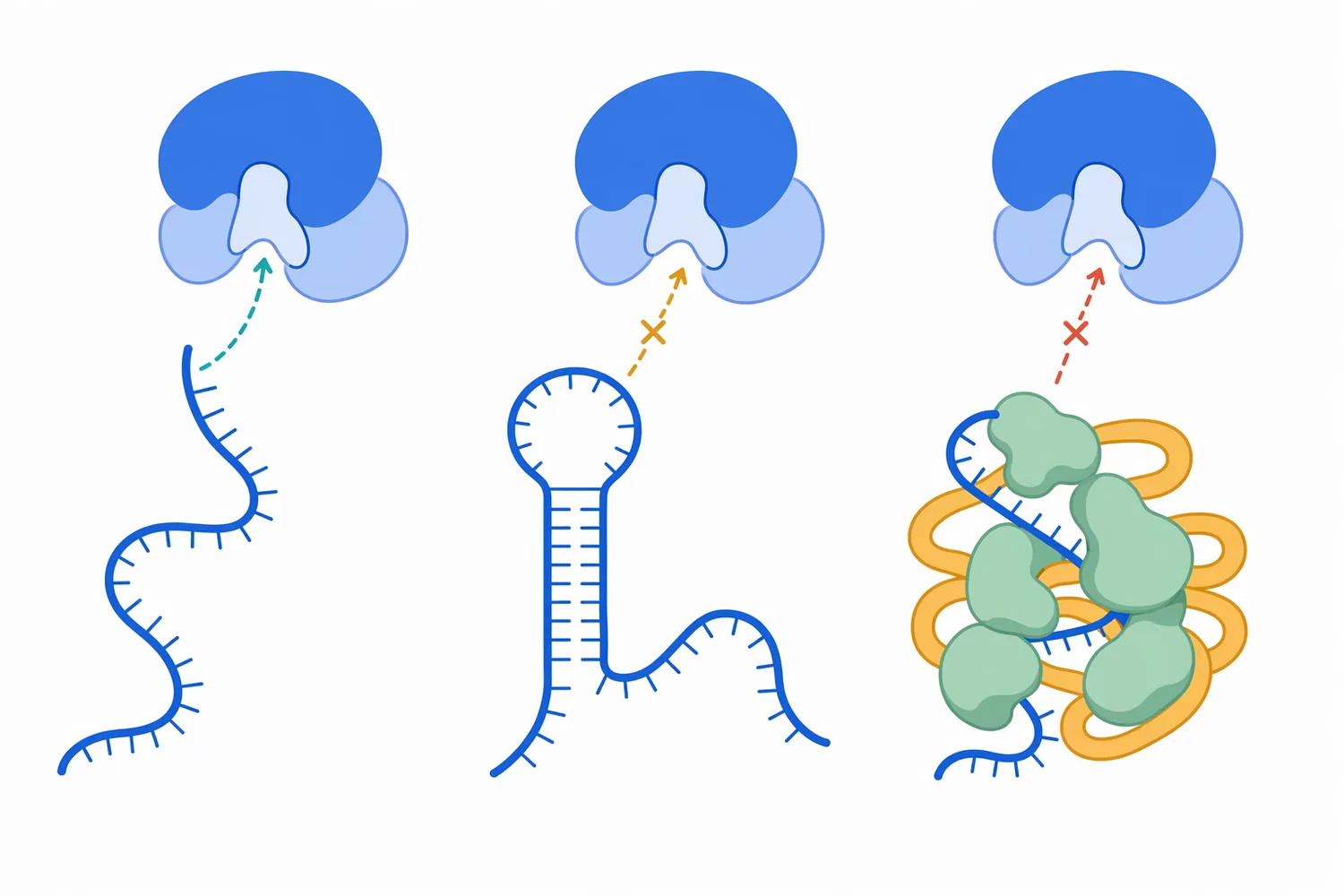
RNase 與 DNase、核酸外切酶、限制酶或聚合酶不同,主要差異在於底物與目的。RNase 的核心目的通常是處理 RNA;DNase 處理 DNA;限制酶辨識特定 DNA 序列;聚合酶則合成核酸。若流程目標是保留 DNA 並移除 RNA,RNase A 類型通常較符合邏輯;若目標是拆解 RNA/DNA hybrid,RNase H 才是更貼近底物的選擇 [1]。
| 酵素類別 | 主要作用對象 | 常見目的 | 與 RNase 的差異 |
|---|---|---|---|
| RNase | RNA 或 RNA 相關結構 | RNA 去除、RNA 加工研究、RNA 結構處理 | 以 RNA 為核心底物 |
| DNase | DNA | 降低 DNA 污染或黏度 | 不適合保留 DNA 的流程 |
| 限制酶 | 特定 DNA 序列 | DNA 剪切與克隆 | 依序列辨識 DNA,不用於一般 RNA 去除 |
| 聚合酶 | 核酸模板與核苷酸 | DNA/RNA 合成 | 催化合成而非降解 |
| 核酸外切酶 | 核酸末端 | 端點修飾、降解或修剪 | 作用方向與底物型態較特定 |
這種比較對製程設計很重要,因為「核酸相關酵素」不是可互換的同義詞。若錯把 RNase 當作一般核酸清除工具,可能在需要保留 RNA 的流程造成失敗;若把 DNase 用在需要保留 DNA 的樣品中,也會破壞目標分析物。酵素的價值來自其專一性,而不是單純的分解能力 [7]。
Enzymes.bio 是酵素供應平台/經銷商,不是 ribonuclease 製造商,也不是檢測實驗室。平台提供的 ribonuclease 產品以 1 kg 單位在線上直接銷售;CoA 與 SDS 會隨訂單一併提供,用於協助買方掌握該批次文件、安全資訊與基本品質資訊。本文不提供活性單位數值、等級宣稱、分析方法或活性單位定義,因為這些資訊應以實際產品文件為準。
對企業使用者而言,RNase 的導入價值在於能以明確機制處理 RNA 問題:RNase A 常用於一般 RNA 去除與模型研究,RNase H 適合 RNA/DNA hybrid,RNase P 與 RNase III 則多見於 RNA 加工與結構性 RNA 研究。實際應用時,應把 RNase 視為製程或研究流程中的功能性工具,並依自身用途、下游分析與合規要求安排內部驗證。
Ribonuclease 是一群功能多元但機制明確的酵素,其共同核心是催化 RNA 或 RNA 相關結構的切割。對 B2B 研發與製程使用者而言,最重要的不是記住所有 RNase 名稱,而是釐清目標底物:是單股 RNA、RNA/DNA hybrid、前驅 tRNA、雙股 RNA,還是與宿主防禦相關的 RNA/膜交互作用。只有把 ribonuclease structure and function、底物特異性與後段流程放在一起評估,RNase 才能在核酸處理、樣品前處理與製程支援中發揮可預期的技術價值 [1]。
以 1 kg 單位販售,現貨供應,可立即出貨。請直接於我們的線上商店下單並付款,我們將為您處理訂單。每筆訂單皆附分析證明書與安全資料表。
購買 Ribonuclease →依首次引用順序編號。所有來源皆為開放取用資料,並於發布時確認可連線;正文中的引用編號會連結至此。