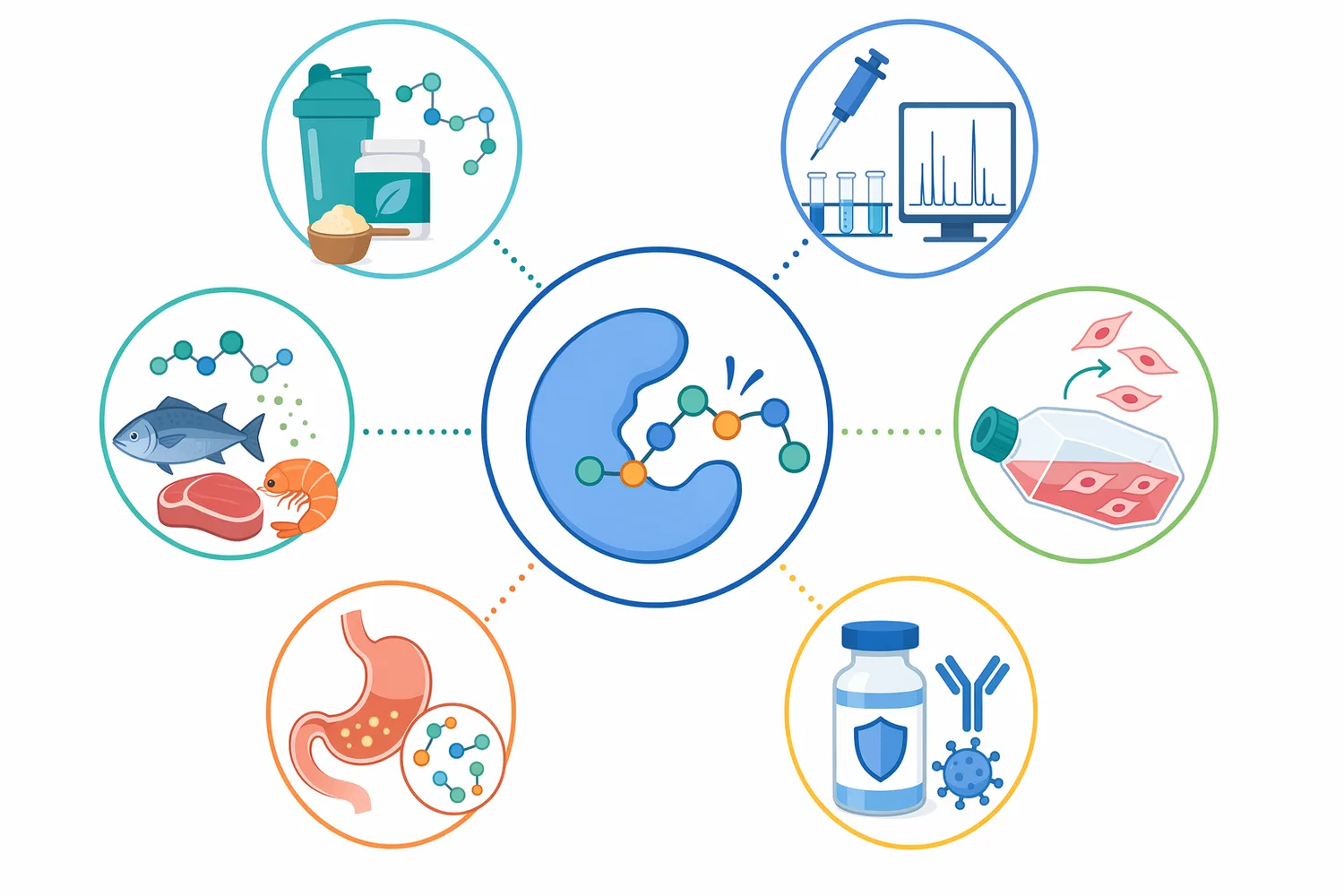
Trypsin is a serine protease used to digest proteins into smaller peptides, with a strong preference for cutting peptide bonds on the carboxyl side of lysine and arginine residues. That defined cleavage pattern is the core trypsin function: it turns large, folded or insoluble protein materials into more manageable peptide mixtures for analytical, biotechnology, food-ingredient, and cell-culture-related workflows [1].
Enzymes.bio supplies Trypsin directly online by the 1 kg unit. Buyers can place and pay for the order online; the order is then processed and shipped, with a Certificate of Analysis and Safety Data Sheet provided with the order.
Trypsin is one of the best-known proteolytic enzymes. In practical terms, a proteolytic enzyme is a catalyst that breaks peptide bonds—the chemical links that hold amino acids together in proteins. The role of trypsin in digestion is to continue the breakdown of dietary proteins after they enter the small intestine, but the same chemistry is valuable outside the body wherever controlled protein hydrolysis is needed [1].
The phrase “what is trypsin?” is often answered too generally as “a digestive enzyme.” For process use, the more useful answer is that trypsin is a substrate-selective serine protease. It does not simply destroy proteins at random. It recognizes peptide-chain regions containing basic amino acids, especially lysine and arginine, and hydrolyzes the peptide bond immediately after those residues when the substrate conformation allows access [1].
This specificity explains why trypsin enzymes are used so broadly in biotechnology and analytical work. A broad protease may reduce a protein to a difficult-to-control mixture; trypsin digestion of protein, by contrast, tends to generate fragments that are more predictable because the enzyme follows a recognizable sequence preference. That predictability is especially valuable in peptide mapping, proteomics, and controlled protein modification workflows [1].
Trypsin is also a member of the larger serine protease family, a group whose catalytic chemistry depends on a highly reactive serine residue in the active site. Modern trypsin research continues to focus on structure, substrate recognition, stability, immobilization, and engineered variants because small changes in the active-site environment or protein surface can alter performance in applied systems [2].
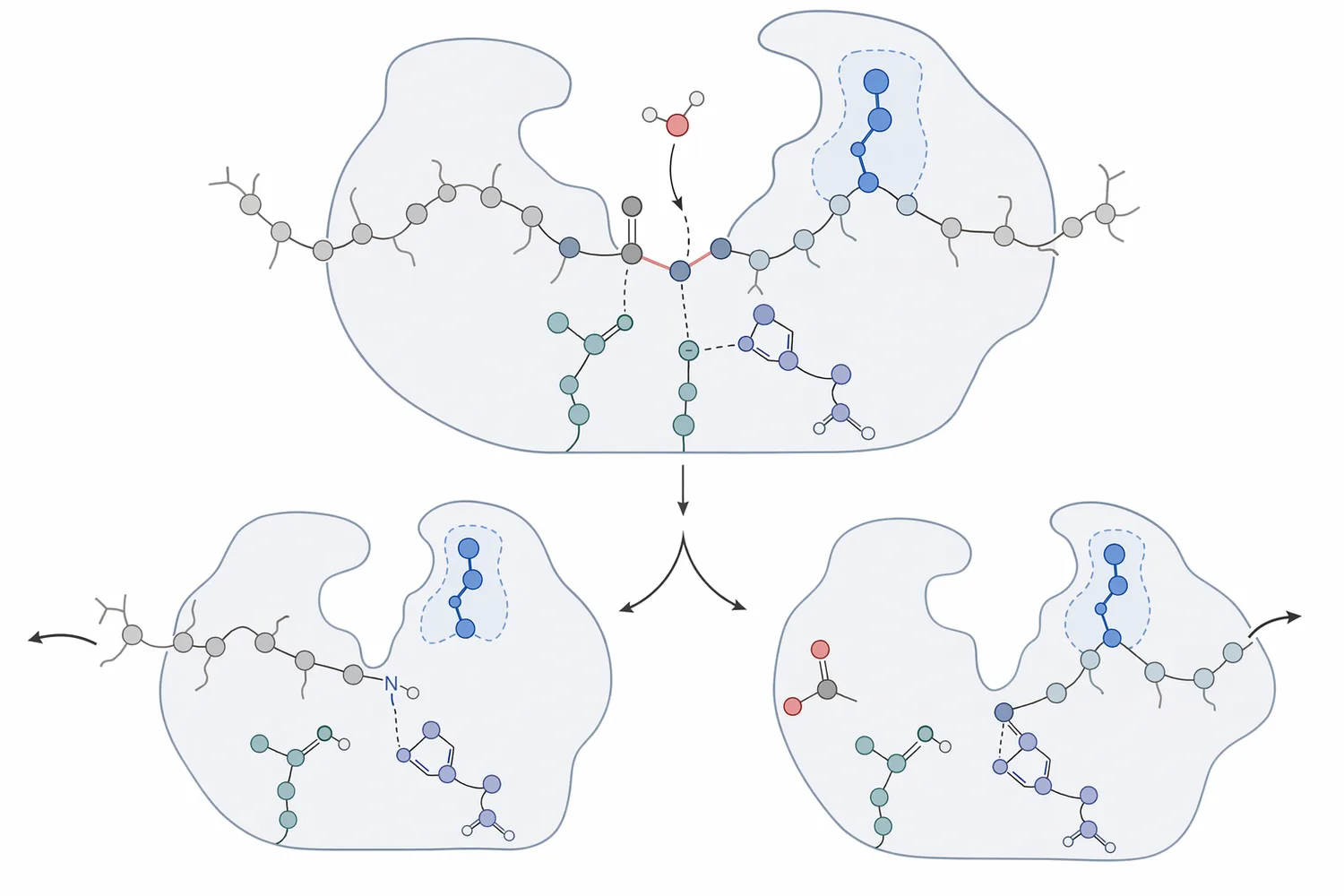
The function of trypsin is peptide-bond hydrolysis. A protein chain contains many peptide bonds, but trypsin preferentially acts where lysine or arginine contributes the amino-acid side chain just before the bond being cut. In simple language, the enzyme binds a protein segment, positions a target peptide bond in the active site, and uses water to split that bond into two shorter peptide chains [1].
The reason trypsin prefers lysine and arginine is structural. Its substrate-binding pocket is suited to positively charged basic side chains, so regions containing those residues fit productively into the enzyme’s recognition site. When the peptide bond is correctly aligned, the catalytic serine attacks the carbonyl carbon of the bond, forming a short-lived intermediate that is then resolved by hydrolysis, releasing peptide products [2].
This is why the trypsin enzyme is often described as a molecular scissors, but the analogy is only partly complete. A pair of scissors cuts wherever the user places it; trypsin cuts only where the protein sequence, local folding, and process environment allow the target bond to enter the catalytic site. In a tightly folded protein, some lysine and arginine sites may be buried and hydrolyze slowly until unfolding, prior cleavage, or process conditions expose them [1].
The visible process outcome is a shift from high-molecular-weight protein to lower-molecular-weight peptides. Depending on the substrate, this can improve solubility, reduce viscosity, expose new functional groups, release bioactive or flavor-active peptide sequences, or create peptide fragments suitable for mass spectrometry. The chemistry is the same across applications; the value comes from what those new peptide products make possible [3].
Trypsin is often discussed alongside pepsin and chymotrypsin because all three digest proteins, but they are not interchangeable. The practical difference is that each enzyme recognizes different substrate features and is associated with a different digestion environment. Understanding pepsin vs trypsin, or trypsin vs pepsin, helps clarify why trypsin is preferred when basic-residue cleavage and predictable peptide generation are desired [1].
| Protease | Conceptual operating environment | Main cleavage tendency | Practical implication |
|---|---|---|---|
| Pepsin | Acidic protein digestion environment | Broad preference for peptide bonds near hydrophobic/aromatic residues | Useful where acid proteolysis is desired, but peptide patterns are generally less trypsin-like and less tied to lysine/arginine cleavage |
| Trypsin | Neutral to mildly alkaline protein digestion environment | Cleaves mainly after lysine and arginine when sites are accessible | Produces comparatively predictable peptide fragments; widely used for controlled digestion and proteomics |
| Chymotrypsin | Neutral to mildly alkaline protein digestion environment | Prefers bonds near bulky hydrophobic or aromatic residues | Complements trypsin by cutting at different sequence positions |
| Trypsin/chymotrypsin combinations | Mixed proteolytic digestion systems | Combines basic-residue and hydrophobic/aromatic cleavage behavior | Can increase hydrolysis breadth when a single enzyme’s specificity is too narrow |
The trypsin/chymotrypsin relationship is especially important in protein digestion. Chymotrypsin and trypsin are both serine proteases, but they “read” protein sequences differently. Trypsin favors lysine and arginine positions, while chymotrypsin favors different side-chain chemistry; together, they can produce a broader digestion profile than either enzyme alone [1].
For buyers comparing proteases, the key point is not that one is universally “stronger.” The key point is substrate fit. If the desired result is cleavage at basic amino-acid residues and reproducible peptide generation, trypsin is the relevant enzyme. If the intended hydrolysis pattern depends on acid digestion or hydrophobic-residue cleavage, pepsin or chymotrypsin may behave differently [1].
When trypsin contacts a protein substrate under suitable aqueous conditions, the first change is molecular rather than visual: peptide bonds begin to break. Each cleavage event converts one long chain into two shorter chains, increasing the number of peptide ends and exposing new amino and carboxyl groups. As digestion progresses, the protein population shifts from intact protein to partial fragments and then to smaller peptides [4].
For soluble proteins, this can reduce molecular size while maintaining a liquid process stream. For insoluble or aggregated proteins, the effect depends on whether trypsin can reach the cleavage sites. Surface-exposed lysine and arginine residues may be cut first, loosening the structure and making additional sites available. In this way, hydrolysis can become progressive: early cuts open the substrate, and later cuts deepen the digestion [1].
For protein-rich raw materials such as plant meals, animal proteins, blood proteins, or specialty protein extracts, trypsin can change functional properties. Hydrolysis may increase dispersibility, alter foaming or emulsifying behavior, reduce gel strength, release peptides with new taste profiles, or create a peptide-rich ingredient stream. The exact result depends on the original protein architecture and how far hydrolysis proceeds [3].
Studies using immobilized trypsin to degrade bovine hemoglobin illustrate the practical concept: the enzyme acts on a large protein substrate and converts it into smaller fragments, while immobilization affects how the enzyme is retained and reused in the system. The central chemistry remains peptide-bond cleavage, but the process format changes how enzyme contact and stability are managed [4].
Trypsin is deeply established in proteomics because proteins are often too large and complex for direct sequence-level analysis. In a typical analytical concept, proteins are digested into peptides, the peptides are separated and measured, and the resulting mass patterns are matched to sequence databases. Trypsin’s lysine/arginine specificity helps generate peptide sets that are interpretable and reproducible [1].
This is one of the clearest examples of the practical trypsin function. The enzyme converts an analytical problem—large, diverse proteins—into a more manageable peptide problem. Because the cleavage rules are known, software and database workflows can predict possible tryptic peptides and compare them with measured signals. That makes trypsin valuable not just as a digestion reagent, but as part of the logic of protein identification [1].
Trypsin also supports peptide mapping in bioprocess and protein-characterization contexts. When a known protein is digested, the resulting peptide pattern can help confirm identity, reveal sequence coverage, or indicate modifications that change peptide mass or detectability. The method depends on consistent digestion behavior, which is why trypsin’s specificity is more useful than a nonspecific protease for many mapping workflows [1].
Research on engineered trypsin variants shows why the enzyme remains an active development area. Semi-rational design and molecular simulations have been used to study how structural changes influence catalytic behavior, providing mechanistic insight into how substrate binding, active-site geometry, and protein dynamics affect trypsin performance [2].
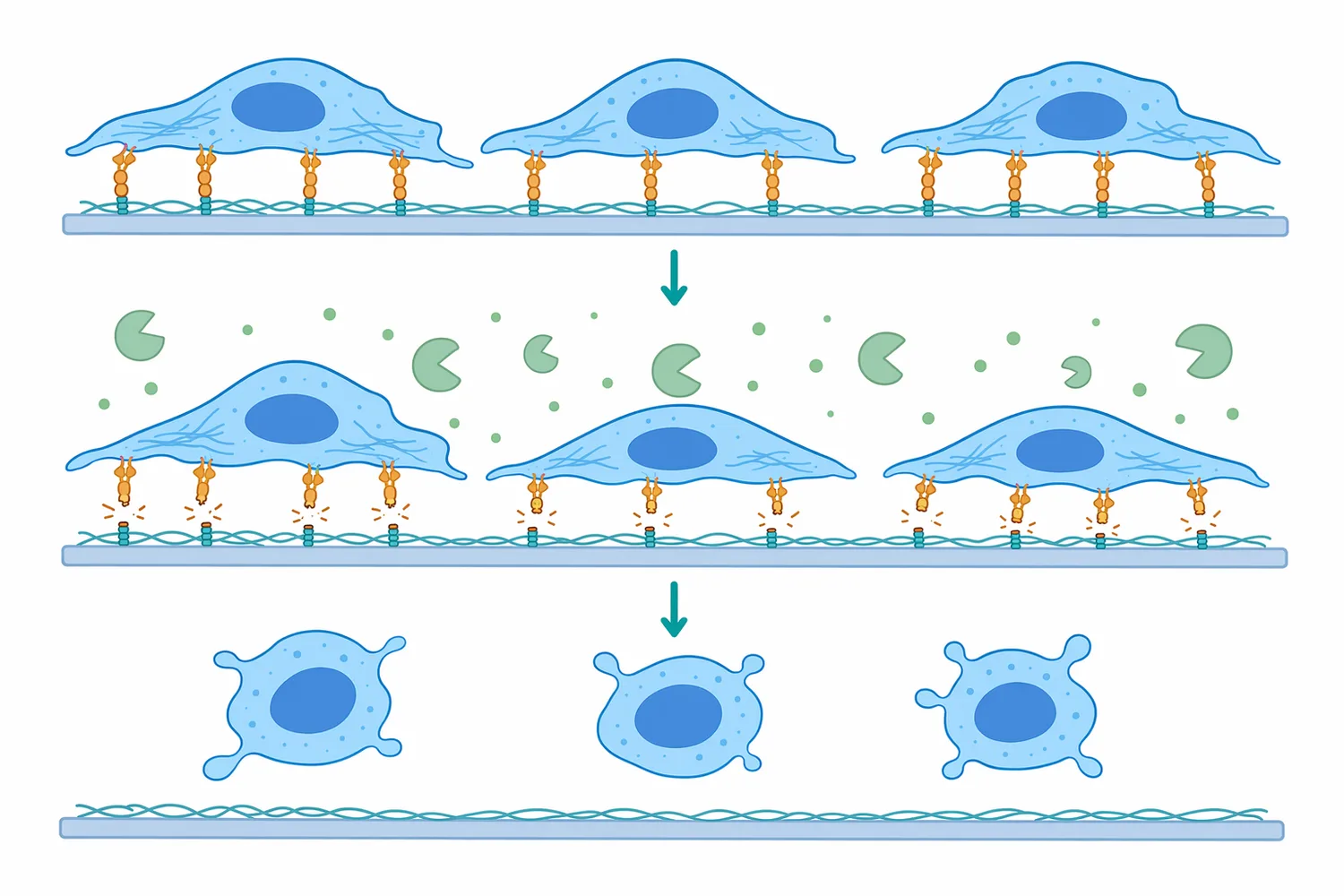
In cell culture, trypsin is commonly associated with detaching adherent cells from growth surfaces. Many adherent cells remain attached through proteins involved in cell-surface and extracellular-matrix interactions. Trypsin can cleave accessible adhesion-related proteins, weakening those interactions so cells can be lifted, resuspended, counted, transferred, or processed further [5].
The mechanism is straightforward: trypsin does not “push” cells off a surface. It cuts protein components that help hold the cells in place. Once enough attachment points are disrupted, normal handling can release the cells into suspension. This makes trypsin useful in routine passaging and scale-up concepts for adherent-cell workflows [5].
That same proteolytic action also explains the caution around sensitive cell types. If the workflow depends on preserving particular surface proteins, receptors, or markers, trypsin exposure can change what remains on the cell surface. Stem-cell culture literature has treated cell dissociation as an important process variable because passaging conditions can influence cell maintenance and downstream handling [6].
Trypsin is also relevant in virus-production contexts where proteolytic processing can influence viral protein activation or cell-culture production systems. Suspension cell lines and scalable culture formats have been studied for influenza virus production, showing how cell-culture engineering and protease-related processing can intersect in applied biotechnology [7].
Food and ingredient applications use trypsin for the same fundamental reason: many food materials are protein-rich, and controlled proteolysis changes how those proteins behave. Hydrolysis can make a protein material more soluble, reduce large insoluble fractions, release shorter peptides, or generate taste-active compounds. In protein hydrolysate production, trypsin helps convert intact proteins into peptide mixtures that can be easier to formulate or further process [3].
Plant proteins are a good example. Fenugreek and ginger proteins have been hydrolyzed using trypsin-based systems to generate peptides investigated for antimicrobial effects, demonstrating how enzymatic digestion can turn raw protein substrates into biologically active peptide fractions. The relevant industrial lesson is that peptide function comes from the sequence and size distribution created during hydrolysis [3].
Animal proteins can also be substrates for trypsin hydrolysis. Bovine hemoglobin degradation by immobilized trypsin has been studied as a model of protein breakdown, showing how a large heme-containing protein can be enzymatically fragmented. This kind of substrate conversion is relevant to broader protein valorization concepts, where the goal is to turn concentrated protein streams into smaller peptide products [4].
Marine and fish-processing applications are another important area in the broader trypsin literature. Atlantic cod trypsins have been studied from basic research through practical applications, including interest in enzymes adapted to cold-water organisms. Such work matters because enzyme source and structure can influence catalytic behavior under different processing environments [8].
Trypsin can also be part of combined protease strategies. In many real protein substrates, one enzyme’s preferred cleavage sites may not be enough to reach the desired hydrolysis profile. A chymotrypsin-trypsin or trypsin chymotrypsin system can broaden the set of cleaved peptide bonds, because each enzyme recognizes different amino-acid environments [1].
Trypsin may be used in soluble form or immobilized on a carrier. Immobilization means the enzyme is attached to, entrapped in, or associated with a solid or structured material so that it can act on substrates while being retained more easily. The benefit is not a change in the core chemistry; the benefit is process handling, enzyme localization, and in some systems improved stability [9].
A Langmuir–Blodgett film system containing immobilized trypsin has been studied for sustained enzymatic activity and rapid detection of azocasein, a protein substrate used to demonstrate proteolytic activity. This illustrates a broader principle: when trypsin is immobilized in an organized matrix, substrate access, enzyme orientation, and local microenvironment can affect observed performance [9].
Other research has examined cuttlebone-modified chitosan as an immobilized trypsin carrier for bovine hemoglobin degradation. In that type of system, the carrier is not the catalyst; trypsin remains responsible for peptide-bond hydrolysis. The carrier changes how the enzyme is presented, retained, and contacted with the protein substrate [4].
Immobilized trypsin is particularly relevant where repeated contact with protein substrates is useful or where enzyme removal from the product stream is desirable. The practical trade-off is that immobilization can improve handling but may also introduce diffusion limits: large proteins must reach the enzyme surface, and not every cleavage site will be equally accessible [9].
Trypsin is itself a protein, which means it can lose activity if its structure is disrupted. Heat, unfavorable chemical environments, oxidation, surface adsorption, or strong inhibitors can alter the shape of the active site and reduce catalytic function. Modern studies using spectroscopy and molecular modeling show that interactions with nanomaterials can change trypsin conformation and enzymatic activity, reinforcing the link between structure and function [10].
Trypsin can also digest itself, a phenomenon known as autolysis or self-digestion. Classic biochemical work examined the self-digestion of trypsin, calcium-trypsin, and acetyltrypsin, showing that trypsin stability is not only about the external substrate but also about the enzyme’s own susceptibility to proteolytic cleavage [11].
Calcium has long been studied in relation to trypsin behavior. Early work on the interaction between calcium and trypsin helped establish that calcium can influence trypsin stability and self-digestion behavior. For applied users, the practical concept is that the enzyme’s structural environment can affect how long it remains active during protein hydrolysis [12].
Surface interactions also matter. Trypsin exposed to graphene oxide–silver nanoparticle systems has been studied for changes in adsorption behavior, conformation, and enzymatic activity. The mechanistic point is that when trypsin binds to a surface, the orientation and flexibility of the protein can change, which may either preserve or impair access to the active site [13].
A trypsin inhibitor protein is a protein or peptide that binds trypsin and reduces its ability to hydrolyze substrates. These inhibitors are common in biological materials, especially seeds and legumes, where they can serve protective roles by interfering with digestive proteases. In processing terms, they can reduce effective trypsin activity if they remain active in the substrate matrix [14].
Soybeans contain well-known inhibitor families, including Kunitz and Bowman–Birk inhibitors. Recent work has examined methods to determine their contributions to total trypsin or chymotrypsin inhibition in soybeans, highlighting that different inhibitor types can contribute differently to overall protease inhibition [14].
The mechanism is direct: an inhibitor protein binds near or within the enzyme’s substrate-recognition region and prevents productive access to the active site. Instead of the protein substrate entering and being cleaved, the inhibitor occupies the enzyme in a nonproductive or slowly reversible complex. The result is slower hydrolysis, incomplete digestion, or the need for upstream processing that changes inhibitor availability [14].
This matters especially when working with plant protein materials. A soybean meal, legume flour, or seed extract may contain both desired protein substrate and protease inhibitors. Trypsin may therefore encounter two competing protein classes: proteins intended for hydrolysis and inhibitor proteins that suppress enzyme function [14].
Trypsin remains important not only because it is old and familiar, but because it is still being improved and adapted. Recombinant expression of trypsin from Streptomyces griseus in Pichia pastoris has been reported, reflecting interest in producing microbial trypsins and trypsin-like enzymes through controlled expression systems [15].
Protein engineering studies continue to investigate how catalytic efficiency, stability, and substrate interaction can be modified. In one recent example, semi-rational design was used to enhance catalytic activity of a novel trypsin, with molecular simulations providing insight into how structural changes affected enzyme behavior [2].
Source diversity is also relevant. Trypsins from different organisms can differ in temperature adaptation, stability, and catalytic characteristics. Atlantic cod trypsins, for example, have attracted attention because cold-adapted marine enzymes may perform differently from mammalian enzymes under lower-temperature processing conditions [8].
These studies do not mean every application needs an engineered or unusual trypsin. They show why trypsin is a robust enzyme platform: its mechanism is well understood, its industrial and analytical uses are established, and its structure can be studied in enough detail to connect molecular changes with process behavior [1].
The most direct use of trypsin is controlled hydrolysis of protein substrates. This includes converting large proteins into peptides for research, ingredient development, analytical preparation, or downstream processing. The value is the combination of proteolytic power and sequence preference: trypsin cuts many proteins effectively, but with a recognizable bias toward lysine and arginine sites [1].
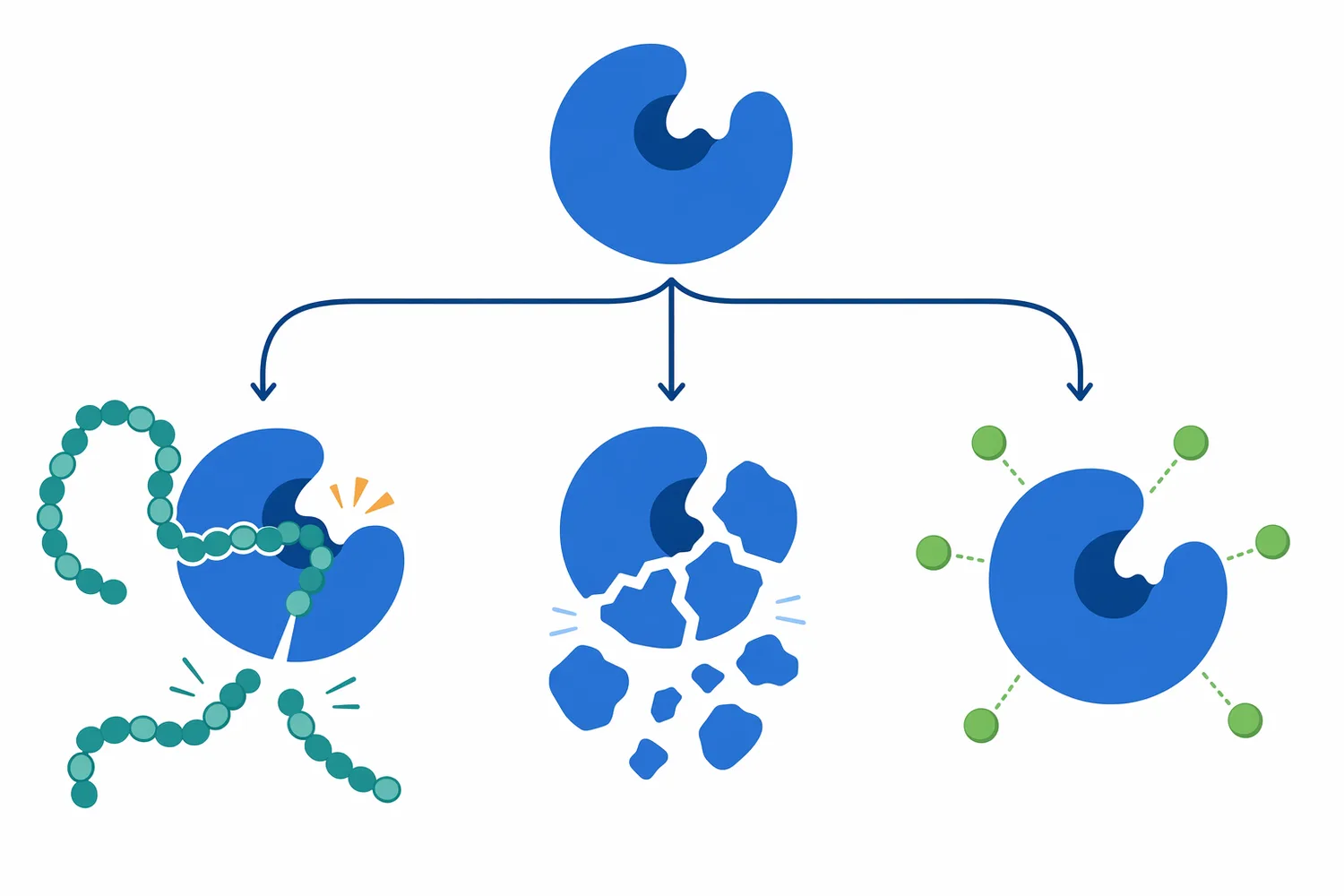
In a hydrolysis process, the visible performance depends on the protein’s structure. A denatured or soluble protein may digest more readily because cleavage sites are exposed. A cross-linked, aggregated, or matrix-embedded protein may digest more slowly because trypsin must first gain access to target bonds. The enzyme’s chemistry is specific, but the substrate’s physical form controls how much of that chemistry is realized [4].
Trypsin can generate peptides with biological or sensory relevance. In fenugreek and ginger protein research, trypsin-based hydrolysis was used to obtain peptides evaluated for antimicrobial activity. This does not mean trypsin itself is an antimicrobial ingredient; it means trypsin can help release peptide sequences whose properties differ from the intact parent protein [3].
For food and ingredient development, this distinction is important. The enzyme is a processing tool. The final peptide mixture depends on the raw protein sequence, the accessibility of cleavage sites, and the extent of hydrolysis. Trypsin changes the molecular composition; the application value comes from the properties of the resulting peptide fraction [3].
For analytical workflows, trypsin’s function is to make proteins measurable as peptides. Large proteins can be difficult to separate, ionize, or identify directly, while tryptic peptides are more compatible with common proteomics approaches. The enzyme therefore acts as a bridge between intact protein complexity and peptide-level analytical readout [1].
This is why trypsin is so closely associated with reproducibility in proteomics. If a protein sequence is known, the likely tryptic fragments can be predicted. If the measured peptides match those predictions, confidence in protein identification increases. Few enzyme-substrate relationships are as deeply integrated into routine protein analysis as trypsin digestion [1].
In adherent cell workflows, trypsin helps release cells by cleaving proteins that mediate attachment. This is a mechanical outcome produced by biochemical action: once enough adhesion proteins are cut, cells no longer remain firmly attached to the culture surface [5].
The same mechanism explains why exposure is controlled in cell-culture practice. Trypsin can affect proteins that are biologically meaningful, not just proteins that attach cells to plastic or extracellular matrix. For sensitive cultures, the balance is between sufficient detachment and unnecessary surface-protein damage [6].
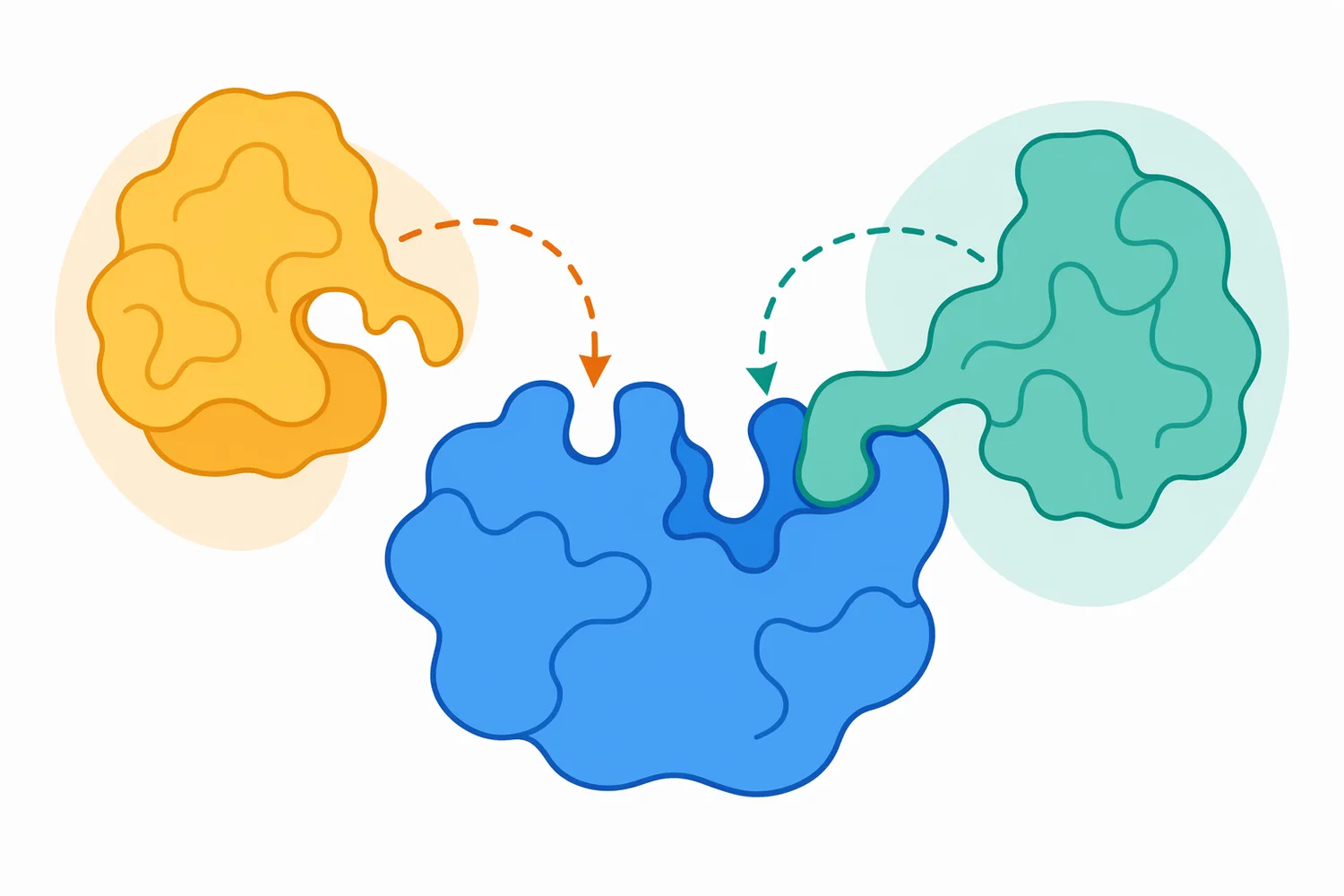
Trypsin is powerful because it is specific enough to be predictable yet broad enough to act on many proteins. That combination makes it useful, but it also means it can modify proteins unintentionally if they are exposed and susceptible. Any protein-containing material in the same environment may become a substrate if the enzyme can access compatible cleavage sites [1].
Performance should be understood as enzyme-plus-substrate behavior, not enzyme behavior alone. The same trypsin may digest one protein rapidly and another slowly because folding, aggregation, inhibitors, salt environment, temperature, surface adsorption, or prior processing change access to cleavage sites. This is why trypsin digestion is best viewed as controlled biochemistry rather than a simple additive effect [10].
Inhibitors are a particularly important reality in natural protein materials. Soybean trypsin inhibitor protein families demonstrate that a substrate can contain compounds that actively suppress trypsin or chymotrypsin. When present, such inhibitors can shift the hydrolysis outcome even if the target protein itself is otherwise suitable [14].
Autolysis is another practical concept. Because trypsin is a protease and also a protein, it can cleave itself under some conditions. Classic studies on trypsin self-digestion and calcium interactions help explain why enzyme stability is tied to the surrounding molecular environment, not just to the initial enzyme preparation [11].
Enzymes.bio supplies Trypsin directly online in 1 kg units for professional users who need a practical enzyme product for controlled protein digestion, peptide generation, research-support workflows, food-ingredient development, or other appropriate protein-processing applications. The purchase process is straightforward: order and pay online, after which the order is processed and shipped.
A Certificate of Analysis and Safety Data Sheet are provided with the order. The product is supplied as a ready-to-purchase enzyme offering rather than through a custom development or laboratory-service model.
Trypsin is best understood as a well-characterized enzyme for converting proteins into smaller peptides with a defined lysine/arginine cleavage preference. Its strongest value is where that predictable proteolysis helps create analyzable, processable, or functionally modified protein materials [1].
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Trypsin →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.