Glucoamylase is a glucose-forming enzyme used to convert liquefied starch and dextrins into fermentable, glucose-rich streams for saccharification, brewing, distilling, ethanol production, and starch-sugar processing. It works mainly from the non-reducing ends of starch-derived chains, removing glucose units step by step after alpha-amylase or heat processing has made the starch more accessible. Enzymes.bio supplies Glucoamylase Starch Saccharification Fermentation Saccharification Enzyme as a 1 kg online product; buyers place the order and pay online, and the order is processed and shipped with a Certificate of Analysis and Safety Data Sheet.
Glucoamylase, also known as amyloglucosidase, is one of the principal enzymes used when the target is glucose rather than partially hydrolyzed dextrins. Starch is made from glucose units arranged mainly as amylose, which is relatively linear, and amylopectin, which is branched. In industrial starch conversion, the starch is normally cooked, gelatinized, liquefied, or otherwise opened up first; glucoamylase then acts on the resulting dextrin chains to release glucose, making it central to the saccharification stage of starch processing [1].
The key distinction is that glucoamylase is not primarily a viscosity-reducing enzyme. Alpha-amylase rapidly cuts internal bonds within gelatinized starch, lowering viscosity and producing shorter dextrins; glucoamylase follows by trimming glucose units from chain ends. This makes it especially useful where the desired output is a fermentable sugar stream for yeast or bacteria, or a glucose-rich syrup rather than a maltodextrin product [2].
For fermentation, the value is direct: many production organisms consume glucose readily, while larger dextrins may remain partly or fully unfermented. In grain mashes, brewing worts, starch hydrolysates, and other saccharified substrates, glucoamylase increases the pool of fermentable sugar by converting residual dextrins into glucose. Studies of starch-rich traditional fermentations also emphasize that the timing and dynamics of starch breakdown determine sugar supply during fermentation, which in turn affects downstream microbial metabolism and product formation [3].
At the molecular level, glucoamylase changes the carbohydrate profile of the process stream. Before saccharification, liquefied starch contains a distribution of dextrins: short and medium-length chains, branched fragments from amylopectin, and smaller oligosaccharides. During glucoamylase treatment, those molecules are progressively shortened from their accessible ends, and free glucose accumulates in the liquid phase [1].
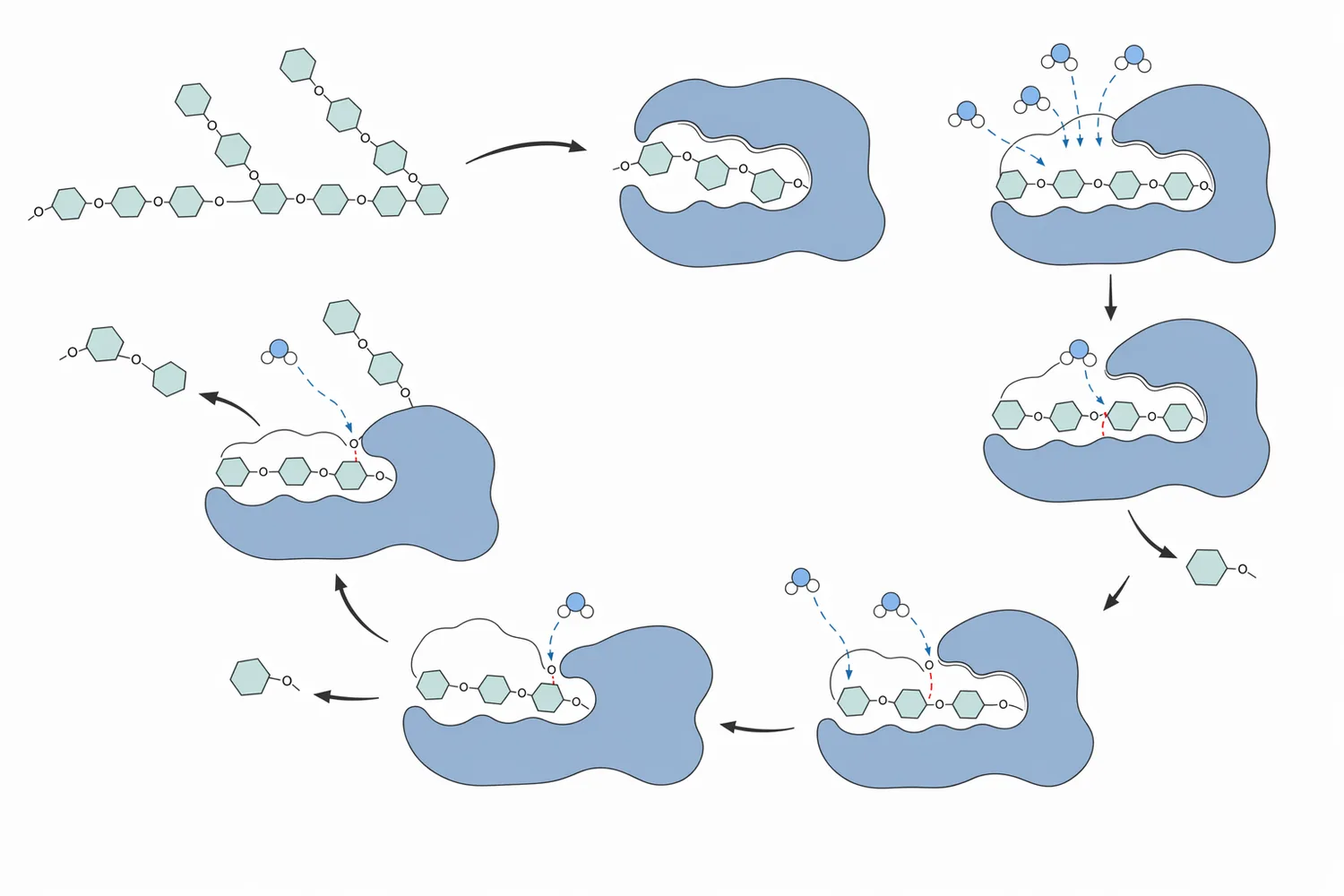
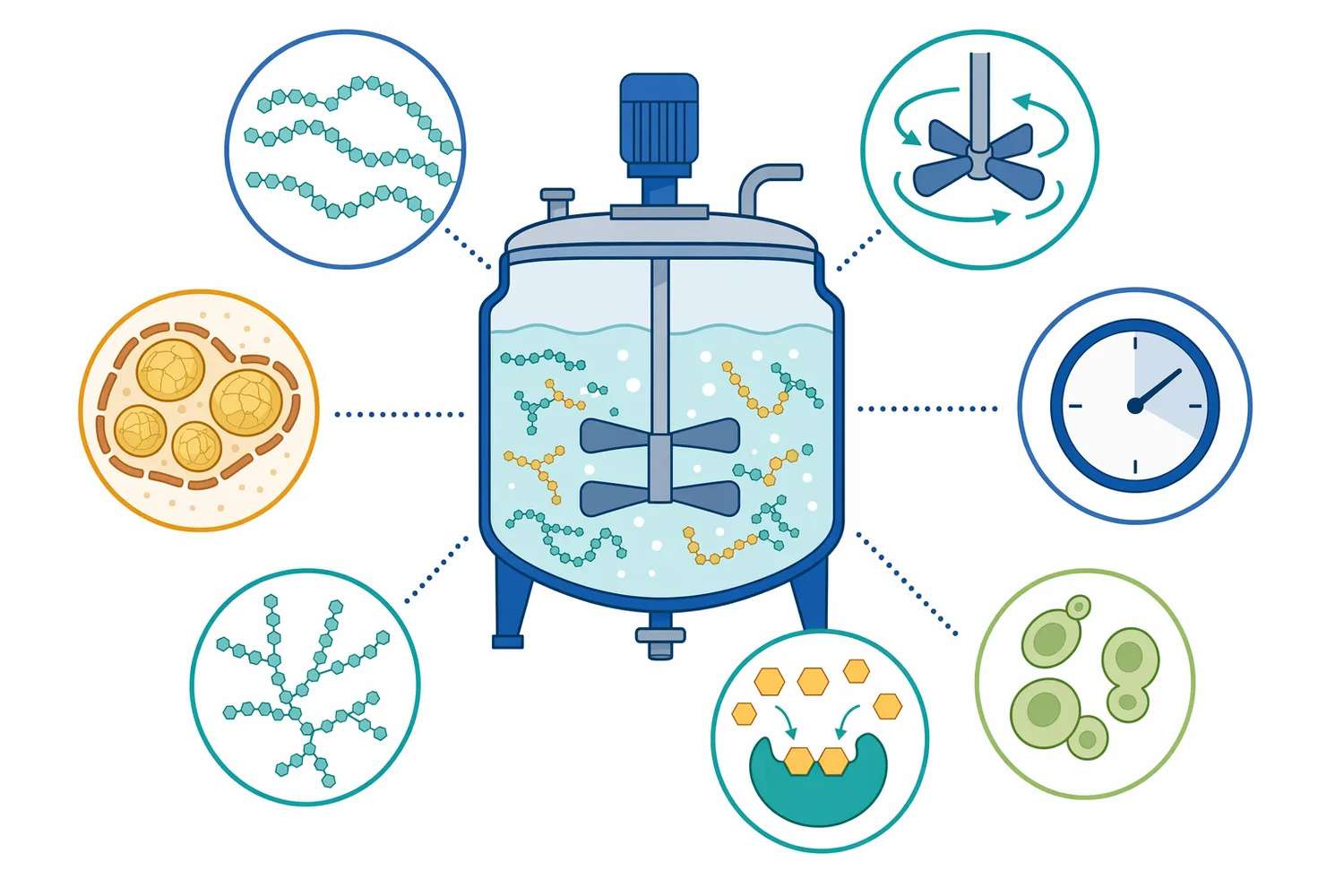
The enzyme’s active site is shaped to bind part of a glucan chain across multiple subsites. These subsites hold glucose residues in the correct orientation so that a terminal glycosidic bond can be hydrolyzed. Research on glucoamylase subsite structure and ligand binding describes this chain-positioning mechanism: the enzyme does not simply “touch starch” in a general way; it recognizes and binds glucose units in a defined arrangement, then cleaves the terminal bond to release glucose [2].
Mutagenesis work on fungal glucoamylase from Aspergillus awamori identified the importance of specific active-site residues, including Asp176, Glu179, and Glu180, in defining the catalytic mechanism. These residues help create the chemical environment needed to break the glycosidic bond: the substrate is held in place, the bond is activated for hydrolysis, and glucose is released as a soluble product [4].
Structural and mechanistic studies have also identified enzyme-substrate and enzyme-product complexes for Aspergillus awamori glucoamylase. This matters for process understanding because it confirms that saccharification is a sequence of binding, cleavage, product release, and rebinding events. The practical result is a gradual shift from dextrin-rich material toward glucose-rich liquor as the enzyme cycles repeatedly through available chain ends [5].
Most starches are not naturally easy for enzymes to access. Native starch granules are semi-crystalline, compact, and protected by granular structure. Heat, moisture, milling, cooking, gelatinization, and alpha-amylase liquefaction all help expose the glucan chains and reduce viscosity so that glucoamylase can work more effectively during the saccharification stage [1].
Liquefaction and saccharification therefore solve different problems. Liquefaction turns a thick starch paste into a pumpable dextrin solution by cutting internal bonds. Saccharification then increases glucose content by working from chain ends. If glucoamylase is used on poorly accessible starch, its reaction may be limited not because the enzyme cannot hydrolyze glucan chains, but because too few suitable chain ends are available or exposed [2].
This is especially important in high-solids processing, grain mashes, and starch-rich fermentation substrates. When viscosity remains high, mixing and mass transfer can limit enzyme contact with substrate. When liquefaction is effective, more dextrin chain ends are exposed in solution, improving the opportunity for glucoamylase to release glucose throughout the batch [1].
Different starch enzymes are useful because they attack different bonds and locations in the starch molecule. The most common conceptual pairing is alpha-amylase for liquefaction and glucoamylase for glucose formation; a debranching enzyme such as pullulanase may also be used where branched amylopectin fragments limit complete conversion [6].
| Enzyme type | Main action on starch-derived material | What changes in the process stream | Typical role in a starch-to-glucose workflow |
|---|---|---|---|
| Alpha-amylase | Cuts internal alpha-1,4 bonds within starch chains | Rapidly lowers viscosity and produces shorter dextrins | Liquefaction before saccharification |
| Glucoamylase | Removes glucose units from accessible chain ends | Increases free glucose and reduces residual dextrins | Saccharification for fermentation or glucose-rich syrup |
| Pullulanase / debranching enzyme | Cleaves alpha-1,6 branch points in amylopectin-derived material | Creates more linear chains and more accessible ends | Supports higher conversion when branched dextrins are limiting |
This difference in action mode is not just theoretical. Research on enzymatically modified starch for paper surface sizing highlights that enzymes with different action modes and sites produce different structural effects in starch. For saccharification, that same principle explains why a process may use one enzyme to open chains, another to release glucose, and another to remove branches when the substrate requires it [6].
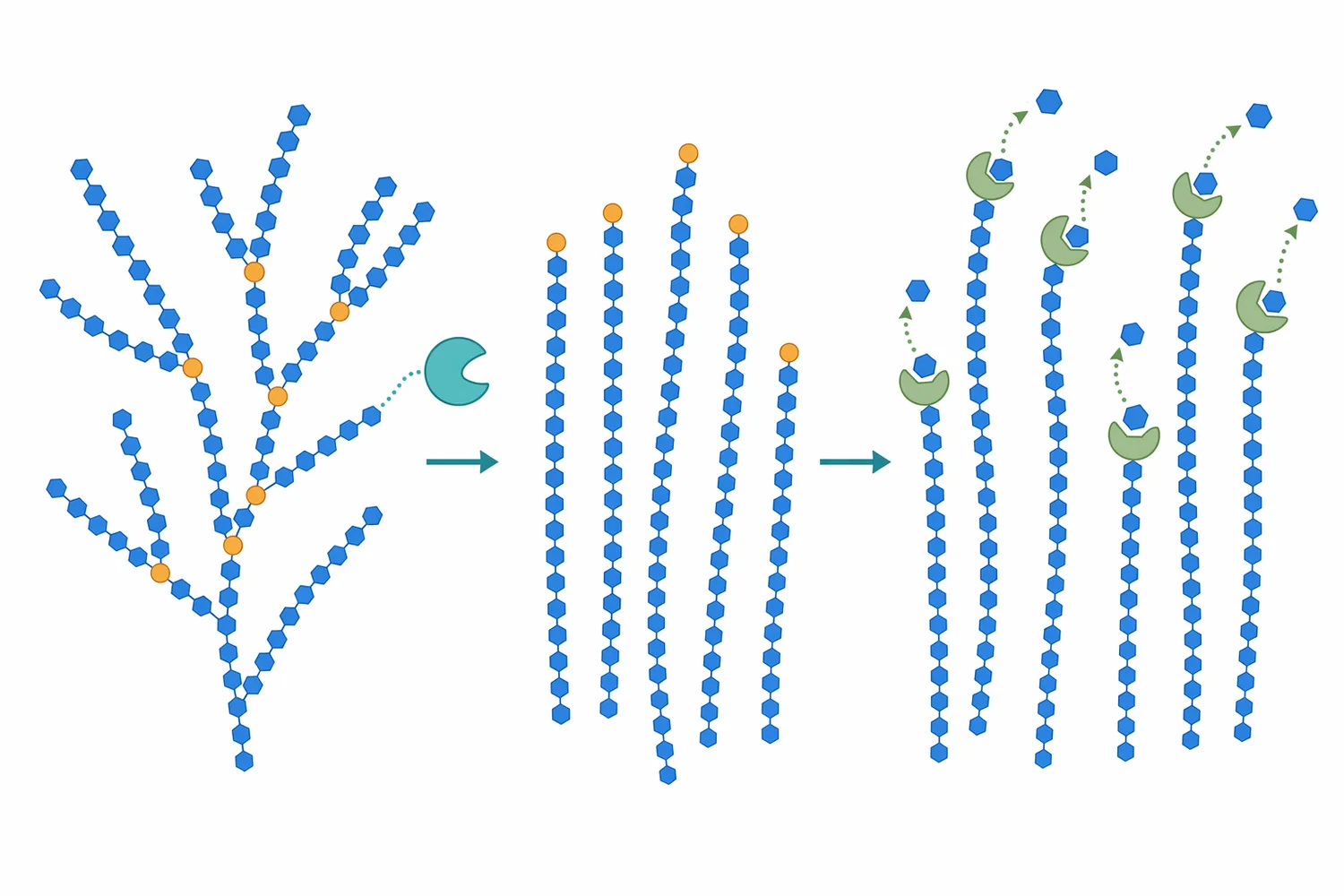
Amylopectin is heavily branched, and those alpha-1,6 branch points can slow complete saccharification. Glucoamylase can act on glucan chain ends and can hydrolyze more than one type of linkage, but branch architecture still affects access and conversion speed. When branches remain, the material may retain dextrin character even after extensive hydrolysis [1].
A debranching step changes the geometry of the substrate. By removing branch points, a debranching enzyme converts a more tree-like amylopectin fragment into more linear chains. Those chains offer additional accessible ends and fewer steric obstacles, which helps glucose-forming enzymes continue saccharification more completely [6].
For a buyer using Glucoamylase Starch Saccharification Fermentation Saccharification Enzyme, the practical takeaway is that glucoamylase is the glucose-releasing component of the saccharification system. Its strongest contribution is converting available dextrins into glucose; when the process stream contains many branched fragments, the broader enzyme system and upstream treatment determine how much substrate is presented in a form that glucoamylase can efficiently finish [2].
The strongest evidence for how glucoamylase works comes from a combination of mutagenesis, substrate-binding studies, and enzyme-product complex analysis. Mutagenesis of Aspergillus awamori glucoamylase at Asp176, Glu179, and Glu180 showed that defined amino-acid residues are required for the enzyme’s catalytic function, supporting a specific active-site mechanism rather than a nonspecific starch-degrading effect [4].
Ligand-binding research adds another layer of explanation. Glucoamylase has subsites that interact with glucose residues in the substrate chain, helping determine how the enzyme binds maltose, longer oligosaccharides, and related glucans. The enzyme’s performance depends on this binding architecture because the terminal bond must be positioned precisely for hydrolysis [2].
Studies identifying enzyme-substrate and enzyme-product complexes help explain why product release matters. Once glucose is formed, it must leave the active site so the enzyme can bind another chain end. In practical saccharification, this repeated turnover is what steadily increases glucose concentration and reduces dextrin length over time [5].
Older mechanistic work comparing glucoamylase action on alpha- and beta-D-glucosyl fluoride also supports the stereochemical specificity of the enzyme’s hydrolytic behavior. That research is useful because it shows that glucoamylase catalysis follows defined carbohydrate chemistry, with substrate orientation and bond stereochemistry influencing the reaction path [7].
Fungal glucoamylases are especially prominent in starch processing because many fungal enzymes are naturally effective at degrading extracellular polysaccharides. Reviews of fungal glucoamylases describe their significance across starch hydrolysis applications, including their ability to release glucose from starch-derived substrates and their long-standing importance in industrial biotechnology [1].
Aspergillus species are among the best-known sources studied for glucoamylase genes and enzymes. Recent work on cloning a glucoamylase gene from Aspergillus niger and expressing it in Pichia pastoris reflects continuing interest in fungal glucoamylase systems and their practical value for starch-converting enzyme supply chains [8].
Research is not limited to Aspergillus. Solid-state fermentation studies with Mucor indicus, Mucor hiemalis, and Rhizopus oryzae have examined glucoamylase production optimization, showing that multiple fungal organisms can produce extracellular glucoamylases relevant to starch degradation [9].
Bacterial glucoamylases have also been studied. Isolation and optimization work on extracellular glucoamylase from Paenibacillus amylolyticus strain NEO03 shows that glucoamylase activity is not exclusive to fungi, although fungal glucoamylases remain especially well established in starch-processing literature [10].
In starch sugar production, the process goal is often to turn corn, wheat, cassava, potato, or another starch source into a glucose-rich hydrolysate. Glucoamylase is the enzyme that drives this shift after the starch has been liquefied into dextrins. The measurable process change is a reduction in dextrin size and an increase in free glucose concentration [1].
This role is valuable because glucose-rich streams are versatile. They can be used directly as sweetener intermediates, fermented into alcohols or organic acids, or further processed into other carbohydrate products. The enzyme’s contribution is not flavor, color, or preservation; it is targeted hydrolysis of starch-derived glycosidic bonds to increase glucose availability [2].
For controlled saccharification, the substrate must be accessible and well dispersed. If starch remains in compact granules or if liquefaction is incomplete, glucoamylase has fewer exposed chain ends to attack. This is why practical starch conversion normally treats glucoamylase as part of a sequence rather than as a single-step solution for every starch material [1].
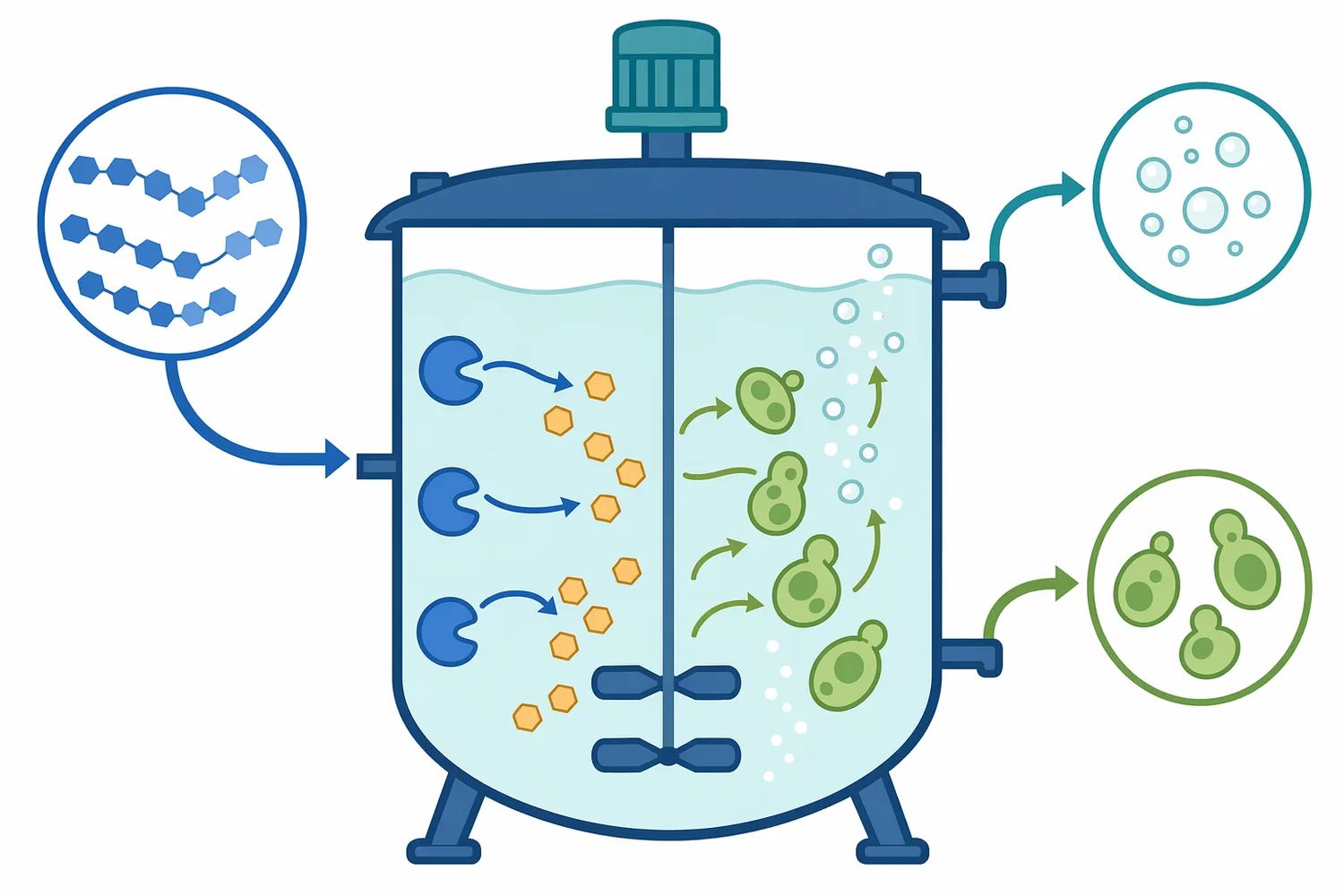
Fermentation performance depends on the type and availability of sugars in the medium. In starch-based systems, starch itself is too large for many production organisms to use directly at industrial rates. Saccharification bridges that gap by generating glucose and smaller fermentable carbohydrates from the original starch material [3].
In beverage and grain fermentations, amylolytic activity is part of the broader transformation from cereal starch into microbial substrate. Reviews of traditional Chinese fermentation starters, such as Qu, describe complex microbial and enzymatic systems that support the conversion of grain components during fermentation, including starch breakdown as part of sugar generation [11].
For ethanol and distilled beverage processes, glucoamylase helps reduce residual dextrins and increase fermentable glucose in mash or wort. The practical effect can be higher substrate utilization when fermentation organisms are glucose-consuming and when other process conditions allow the released sugar to be converted efficiently [1].
In simultaneous saccharification and fermentation concepts, glucose is generated and consumed in the same overall process environment. The advantage is that glucose release can be paired with microbial uptake, potentially reducing sugar accumulation and aligning saccharification with fermentation demand. The suitability of this approach depends on the organism, substrate, process temperature, and the enzyme system being used [3].
In brewing, residual dextrins contribute body, sweetness perception, and final gravity. In highly attenuated beers, dry beer styles, specialty fermentations, or distilling mashes, a higher degree of dextrin conversion may be desirable. Glucoamylase supports that goal by converting dextrins into glucose that yeast can ferment [1].
The same chemistry applies in distilling mashes. Grain starch is first made accessible and hydrolyzed into soluble carbohydrates; glucoamylase then increases the fermentable fraction by releasing glucose. The visible process outcome is not simply “more enzyme action,” but a changed carbohydrate spectrum: fewer larger dextrins and more fermentable sugar [2].
This is why glucoamylase is often discussed alongside attenuation. Attenuation reflects the extent to which fermentable extract is consumed during fermentation, and glucoamylase can increase the fermentable portion of starch-derived extract when dextrins would otherwise remain. The final result still depends on yeast performance, mash composition, and the overall fermentation design [11].
Although glucoamylase is most closely associated with saccharification, its ability to hydrolyze starch from chain ends can also be useful in controlled starch modification. In applications where starch structure, molecular size distribution, or surface properties matter, different enzymes create different physical and functional outcomes [6].
For example, research on enzymatically modified starch for paper surface sizing examines enzymes with different action modes and sites. That work is relevant beyond paper because it demonstrates a broader principle: enzyme choice changes starch architecture in predictable ways. Glucoamylase-type exo-action tends to remove terminal glucose units, while endo-acting enzymes produce more internal cleavage and rapid chain shortening [6].
In food and bioprocess settings, this means glucoamylase can be used when the target is glucose formation or dextrin reduction, not simply starch thinning. Where the desired product is a particular texture, viscosity, or dextrin profile rather than glucose, other amylolytic enzymes may play the dominant role [1].
Glucoamylase performance depends on the state of the substrate. Gelatinized or liquefied starch presents more accessible chains than compact raw granules, and shorter dextrins provide more available chain ends than intact starch. This is why upstream milling, cooking, liquefaction, and mixing affect the apparent success of saccharification [1].
The carbohydrate composition also matters. Amylose-rich material behaves differently from highly branched amylopectin-rich material, and branch points can slow complete conversion. When a process contains many branched dextrins, debranching activity can improve the amount of linear substrate available for glucose release [6].
Process environment matters as well, but it should be understood as part of the full workflow rather than as an isolated enzyme label. pH, temperature, solids content, residence time, mixing, and fermentation compatibility all affect the rate at which dextrins are converted and glucose is consumed or accumulated. Research on enzyme-substrate binding reinforces that saccharification is governed by contact, binding, catalysis, and release, not by enzyme addition alone [2].
Product inhibition and substrate effects can also influence real batches. As glucose accumulates, the reaction environment changes; as dextrins are depleted, the enzyme encounters fewer suitable chain ends. This is one reason industrial saccharification is managed as a time-dependent conversion process rather than an instantaneous treatment [5].
Glucoamylase is highly useful, but it is not a universal fix for every starch-processing problem. If starch is not gelatinized, milled, cooked, or otherwise accessible, the enzyme may not reach enough glycosidic bonds to deliver the expected glucose formation. Raw-starch conversion requires enzymes and process conditions specifically suited to raw granule attack, and that should not be assumed for every glucoamylase application [1].
It also does not replace alpha-amylase in conventional liquefaction. Alpha-amylase reduces viscosity by cutting internal chain bonds, while glucoamylase releases glucose from accessible ends. A process that needs both viscosity reduction and glucose formation usually relies on both action modes, either sequentially or within a broader enzyme system [6].
Glucoamylase also does not remove every structural limitation caused by amylopectin branching. It can contribute to hydrolysis of starch-derived material, but branch points may remain a practical bottleneck in some substrates. In those cases, the overall saccharification system—not glucoamylase alone—determines how completely branched dextrins are converted [2].
Finally, fermentation results are not determined only by sugar release. Yeast or bacterial strain, nutrient balance, inhibitors, osmotic pressure, alcohol tolerance, and process timing all influence final yield and productivity. Glucoamylase improves fermentable sugar availability, but the fermentation organism must still convert that sugar efficiently [3].
The main reason to use glucoamylase is to increase glucose formation from starch-derived dextrins. In a saccharification tank, mash, wort, or hydrolysate, that means shifting carbohydrate material from larger, less fermentable molecules toward glucose. For fermentation, this improves the availability of a sugar that many organisms can metabolize readily [1].
A second reason is consistency. Enzyme-based saccharification gives processors a controlled biochemical route for converting starch, rather than relying only on variable endogenous enzymes from grain or fermentation starters. Traditional fermentations demonstrate the importance of enzyme-driven starch breakdown, but added glucoamylase allows the glucose-forming step to be more deliberately incorporated into a process [11].
A third reason is compatibility with established starch workflows. Glucoamylase fits naturally after liquefaction, alongside other amylolytic enzymes, and within fermentation systems where glucose release is needed. Its mechanism is well studied, its role in fungal starch degradation is well documented, and its application logic is straightforward: expose dextrins, bind chain ends, hydrolyze terminal bonds, and release glucose [4].
Enzymes.bio supplies Glucoamylase Starch Saccharification Fermentation Saccharification Enzyme as an online product for starch saccharification and fermentation-related use. The product is sold directly online by the 1 kg unit: buyers add the product to cart, pay online, and the order is processed and shipped.
A Certificate of Analysis and Safety Data Sheet are included with the order. This keeps the purchasing process simple for users who already know they need a glucoamylase enzyme for starch conversion, mash saccharification, brewing, distilling, ethanol fermentation, or glucose-rich hydrolysate preparation.
For practical purposes, this enzyme is best understood as the glucose-forming component in a starch saccharification workflow. It is used where liquefied starch, dextrins, or starch-derived oligosaccharides need to be converted into a more fermentable or glucose-rich stream through controlled enzymatic hydrolysis [1].
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Glucoamylase 200,000 U/G Starch Saccharification Fermentation Saccharification Enzyme →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.