グルコアミラーゼは、液化後のでんぷん由来デキストリンを非還元末端側から加水分解し、発酵で利用しやすいグルコースへ近づける糖化酵素です。でんぷん糖化では、α-アミラーゼによる液化、グルコアミラーゼによる糖化、必要に応じた脱分岐酵素の補助が工程性能を左右します[1]。
Enzymes.bioは、本グルコアミラーゼを製造業者や研究機関としてではなく、B2B向け酵素供給業者としてオンラインで提供します。製品は1kg単位で直接購入でき、注文時にはCoAおよびSDSが併せて提供されます。
グルコアミラーゼは、でんぷん、マルトデキストリン、短鎖オリゴ糖などをグルコースへ進めるために使われるエキソ型の糖質加水分解酵素です。α-アミラーゼがでんぷん鎖の内部を切って粘度を下げ、液化物をつくるのに対し、グルコアミラーゼはその後段で糖鎖末端からグルコースを切り出す役割を担います。でんぷん糖化を目的とした研究では、α-アミラーゼ、グルコアミラーゼ、プルラナーゼを組み合わせた三酵素系が「一段階のでんぷん加水分解活性」を持つ触媒として検討されており、複数酵素の役割分担が糖化設計の中心であることを示しています[1]。
発酵工程で重要なのは、単にでんぷんを短くすることではなく、酵母や発酵微生物が取り込みやすい糖へ変換することです。グルコースは多くの発酵微生物にとって直接利用しやすい炭素源であり、糖化が不足すると、発酵槽内にデキストリンや分岐オリゴ糖が残りやすくなります。近年の発酵研究でも、でんぷん構造の変化と糖供給の動態が初期発酵の進行に影響することが扱われており、糖化段階は発酵前処理ではなく、発酵性能を決める基盤工程として理解する必要があります[2]。
本製品は、でんぷん糖化、発酵原料調製、醸造・食品加工、糖化液製造などで使われるグルコアミラーゼ系酵素として位置づけられます。ただし、すべての原料を単独で完全糖化する万能酵素ではありません。原料でんぷんの由来、糊化・液化の状態、固形分、pH、温度、滞留時間、共存酵素の有無によって糖組成は変わります。したがって実務上は、「液化後のデキストリンをグルコース側へ押し進める糖化酵素」として設計に組み込むのが最も現実的です[1]。
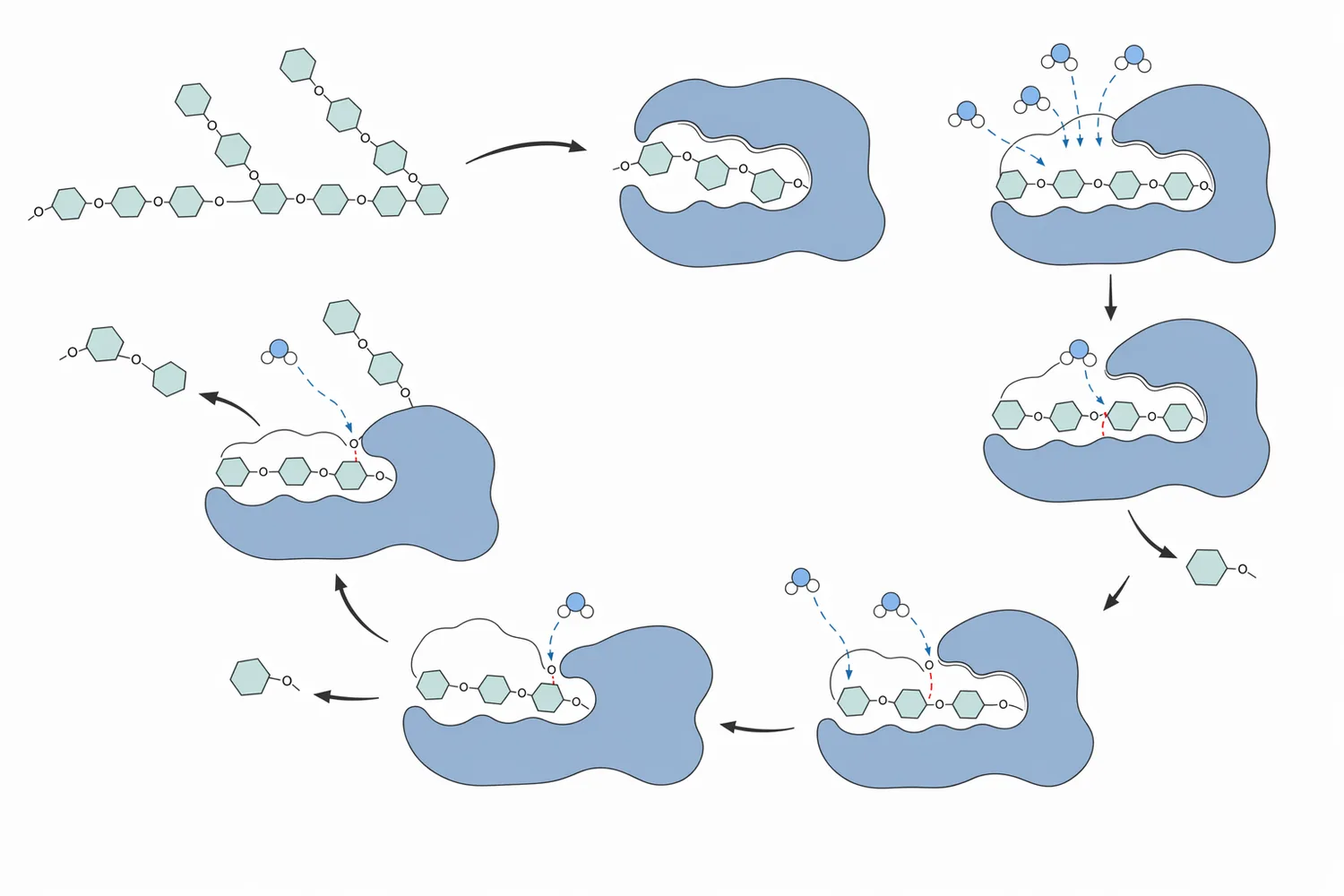
グルコアミラーゼの基本反応は、でんぷん由来糖鎖の非還元末端に作用し、グルコース単位を順に遊離する加水分解です。でんぷんは主に、直鎖に近いアミロースと、多数の分岐を持つアミロペクチンから構成されます。液化後のマルトデキストリンにも、直鎖状部分と分岐点を含む断片が混在します。グルコアミラーゼはこれらの末端を基質として認識し、糖鎖を短くしながらグルコース濃度を高めます。Aspergillus niger由来グルコアミラーゼの脱糖鎖化研究では、酵素の構造、活性、安定性が密接に関連することが示されており、触媒性能は単なる「でんぷん分解能力」ではなく、タンパク質構造の維持と基質認識に依存します[3]。
分岐でんぷんでは、α-1,6結合を含む枝分かれ部分が糖化の進行を制約します。グルコアミラーゼは主として末端側から作用するため、分岐点が多い原料では、脱分岐酵素であるプルラナーゼなどとの組み合わせが理にかないます。α-アミラーゼ、グルコアミラーゼ、プルラナーゼを同時に固定化した研究では、三酵素が一体となってでんぷん加水分解を進める設計が検討されており、液化、末端糖化、脱分岐の三要素が一つの糖化プロセスで相補的に働くことを示しています[1]。
反応の実務的な意味は明確です。α-アミラーゼだけでは、粘度低下や鎖長短縮は進んでも、最終糖組成が十分にグルコースへ寄らない場合があります。一方、グルコアミラーゼだけを未液化のでんぷんに加えても、基質表面や糊化状態が制約となり、期待通りに進まない場合があります。そのため、でんぷん糖化では「液化で基質を開く」「糖化でグルコースを増やす」「必要に応じて脱分岐で残存構造をほどく」という順序で考えると、酵素の役割が整理しやすくなります[1]。
| 酵素 | 主な作用対象 | 工程上の役割 | グルコアミラーゼとの関係 |
|---|---|---|---|
| α-アミラーゼ | でんぷん鎖内部の結合 | 液化、粘度低下、デキストリン生成 | グルコアミラーゼが作用しやすい短鎖基質を増やす |
| グルコアミラーゼ | デキストリンやオリゴ糖の非還元末端 | グルコース生成、発酵可能糖の増加 | 糖化工程の中核酵素 |
| プルラナーゼ | 分岐点のα-1,6結合 | 脱分岐、残存デキストリン低減の補助 | 分岐構造をほどき、グルコアミラーゼの末端作用を補助 |
| 複合酵素系 | 液化物全体 | 糖化速度、糖組成、残存糖の調整 | 三酵素系の研究では、でんぷん加水分解活性の統合が検討されている[1] |
この比較から分かるように、グルコアミラーゼは「でんぷんを最初に壊す酵素」ではなく、「液化後の糖鎖を発酵で使いやすい単糖へ仕上げる酵素」と見る方が正確です。特に高固形分の糖化液、穀物由来原料、醸造用糖化、バイオエタノール原料では、液化の程度がグルコアミラーゼの見かけの働きに強く影響します。前段が不足していると、酵素量を増やしても糖化率が伸びにくいことがあり、逆に液化と脱分岐が適切であれば、グルコアミラーゼの末端加水分解が効率よく進みます[1]。
発酵では、糖の総量だけでなく、糖がいつ、どの形で供給されるかが重要です。発酵初期に利用可能糖が不足すると、微生物の増殖や代謝立ち上がりが遅れます。一方で、糖化液の組成が過度に偏ると、浸透圧、発酵熱、代謝副産物、風味形成に影響します。中国伝統発酵スターターであるQuに関する研究では、発酵スターターの微生物多様性、調製工程、食品応用が整理されており、発酵は単一微生物だけでなく、原料、糖供給、酵素活性、微生物群集が重なり合うプロセスであることが示されています[4]。
グルコアミラーゼは、このうち「でんぷんから糖への供給」を支える要素です。たとえば、穀物発酵では、米、麦、トウモロコシ、芋類などのでんぷんが原料になりますが、微生物が直接利用できるのは主に単糖や一部の二糖です。液化・糖化が適切であれば、発酵開始時に利用可能な糖が増え、発酵工程の再現性を高めやすくなります。でんぷん構造と糖供給動態を扱った発酵研究でも、発酵初期の糖の出方が工程解析の対象になっており、糖化酵素は発酵前の補助剤ではなく、発酵基質の品質を決める要素として扱うべきです[2]。
同時糖化発酵のように、糖化と発酵を重ねて行う考え方もあります。この場合、グルコアミラーゼがデキストリンからグルコースを生成し、生成した糖を微生物が順次消費するため、糖蓄積を抑えながら発酵を進められる可能性があります。ただし、酵素が好む条件と発酵微生物が好む条件は完全には一致しないため、温度やpHは両者の妥協点で設計されます。ここでも、グルコアミラーゼ単体の性能だけでなく、発酵生物、原料処理、工程時間の組み合わせが結果を左右します[4]。
バイオエタノールや発酵化学品では、でんぷん質原料を糖へ変換し、微生物発酵へつなげる工程が基本になります。グルコアミラーゼは、液化済みのでんぷん由来デキストリンをグルコースへ進めることで、酵母などが利用しやすい炭素源を供給します。糖供給が遅い、または不完全な場合、発酵時間の延長、残糖の増加、生成物濃度のばらつきにつながる可能性があります。発酵初期におけるでんぷん構造の変化と糖供給を扱う研究は、糖化段階が発酵性能に直接関係することを示しています[2]。
醸造や発酵食品では、でんぷんをどの程度グルコースへ進めるかが、アルコール生成、酸生成、風味、ボディ感に影響します。グルコアミラーゼを使用すると、発酵可能糖が増えやすくなるため、糖化不足による発酵停滞を避けたい場合に有用です。一方で、すべてを速く単糖化すればよいわけではなく、製品ごとの風味設計や微生物の糖利用順序も考慮されます。伝統的な発酵スターターの研究では、調製工程、微生物多様性、食品応用が密接に結びついており、酵素処理はその一部として理解されます[4]。
でんぷん糖製造では、液化物を糖化し、目的に応じた糖組成へ近づける必要があります。グルコアミラーゼは、グルコース比率を高める方向に作用するため、グルコースシロップや発酵用糖液の製造に向いています。分岐構造が多い場合には、プルラナーゼなどの脱分岐酵素との併用で、残存分岐デキストリンの処理を補助する設計が考えられます。α-アミラーゼ、グルコアミラーゼ、プルラナーゼの複合固定化研究は、こうした役割分担を一つの触媒系にまとめる発想を示しています[1]。
食品・飲料製造では、糖化は単なる収率向上だけでなく、品質の再現性にも関係します。糖組成が変わると、甘味、発酵速度、粘度、褐変反応、最終製品の官能特性が変化します。また、食品・飲料工場ではHACCPや保全活動を統合し、工程リスクを管理する考え方が重視されています。酵素を使う工程でも、投入、保管、記録、安全データの管理を既存の品質・安全管理の流れに組み込むことが実務上重要です[5]。
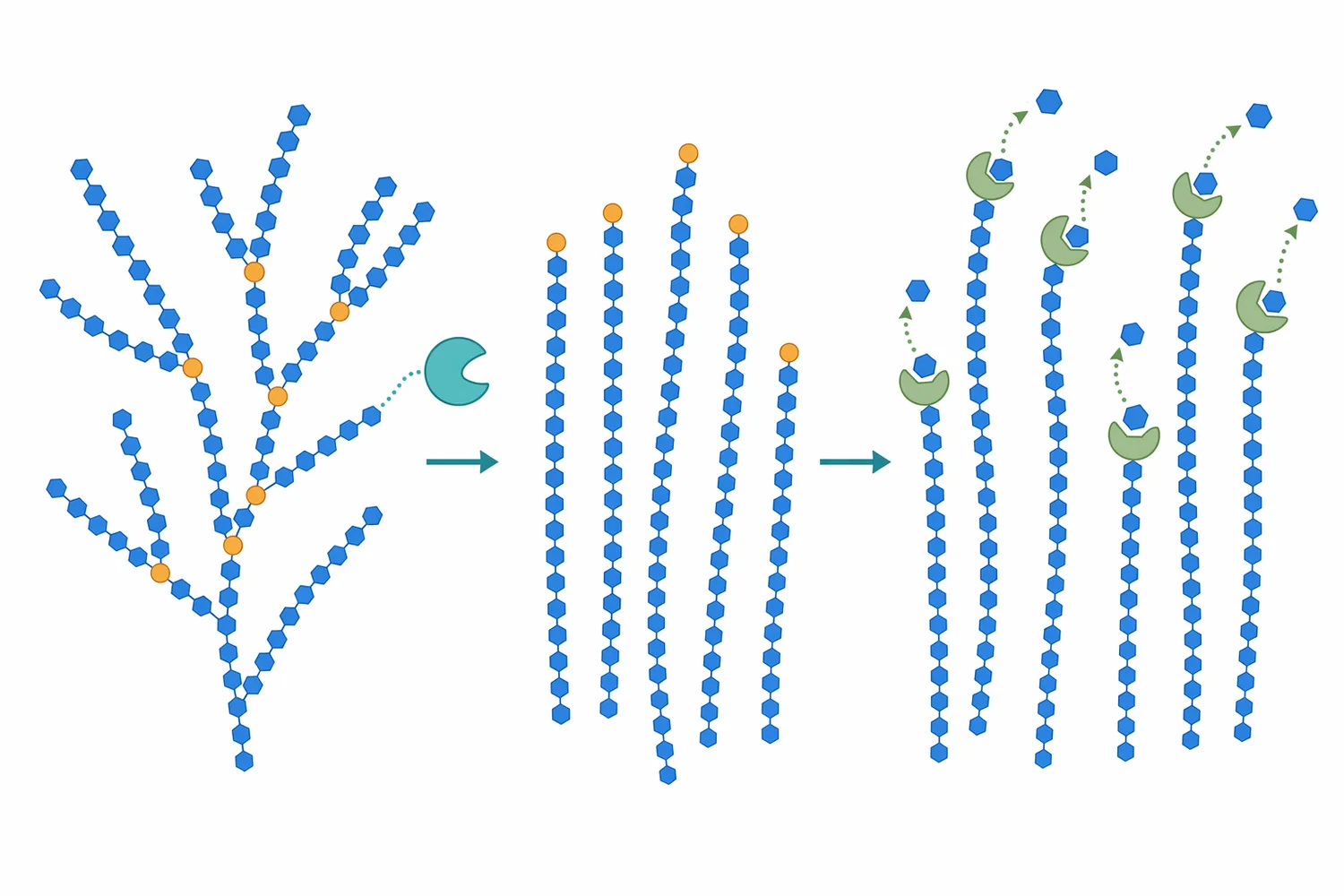
グルコアミラーゼに期待できる第一の効果は、液化後のでんぷん由来糖鎖をグルコースへ進め、発酵可能糖を増やすことです。これにより、発酵原料の利用性が高まり、残存デキストリンを低減しやすくなります。第二の効果は、糖化液の設計自由度です。α-アミラーゼ、グルコアミラーゼ、プルラナーゼを組み合わせることで、粘度低下、末端糖化、脱分岐という異なる反応を工程内で分担できます[1]。
一方で、グルコアミラーゼを加えれば、どの原料でも同じ糖化結果になるわけではありません。原料粒子の状態、糊化の程度、液化の進み方、分岐構造、阻害物質、発酵微生物の糖消費速度が結果に影響します。さらに、酵素タンパク質は熱、pH、界面、金属イオン、糖濃度などの影響を受けます。Aspergillus niger由来グルコアミラーゼの糖鎖除去研究では、糖鎖構造が酵素の構造、活性、安定性に影響することが扱われており、酵素性能は分子構造と工程環境の両方に依存します[3]。
また、生でんぷんを直接処理できるかどうかは、個別の酵素特性に依存します。一般的な工程では、でんぷんを糊化・液化してからグルコアミラーゼで糖化する流れが設計しやすく、未処理原料への直接作用を前提にしない方が安全です。研究領域では、安定性や再利用性を高めるための固定化、ナノ構造化、MOF担持などが盛んに検討されていますが、これらは研究上の酵素設計であり、すべての市販酵素にそのまま当てはまるものではありません[6]。
グルコアミラーゼに関する研究では、活性だけでなく安定性の向上が大きなテーマになっています。生体由来の酵素は高選択的で効率的ですが、熱、pH、せん断、溶媒環境によって構造が変化すると、触媒能が低下します。そのため、酵素を担体に固定化する、架橋凝集体にする、無機・有機ハイブリッド構造へ組み込むといった研究が進められています。グルコアミラーゼを生物学的MOFに組み込む研究では、安定性を高める設計が報告されています[6]。
κ-カラギーナン表面へグルコアミラーゼを共有結合固定化する研究も、酵素安定性と再利用性を改善する方向の一例です。こうした固定化は、酵素分子の自由度を制限し、変性しやすい構造変化を抑えることを狙います。ただし、固定化によって基質拡散が制限される場合もあり、活性、安定性、反応速度、回収性のバランスが重要です。これは市販粉末酵素をそのまま説明するものではなく、グルコアミラーゼが工程条件に敏感なタンパク質触媒であることを理解するための研究知見です[7]。
自己組織化した有機・無機ハイブリッドのグルコアミラーゼナノフラワーや、グルコアミラーゼを埋め込んだMOFに関する研究では、活性や安定性を高める材料設計が報告されています。これらの研究は、産業糖化で求められる「長時間安定に反応する」「温度変化に耐える」「繰り返し利用できる」といった課題に対し、酵素周辺環境を制御するアプローチを示しています[8][9]。
磁性架橋酵素凝集体やFe₃O₄/グラフェン酸化物ナノ複合体への固定化研究も、グルコアミラーゼの安定性と再利用性を高める方向で検討されています。磁性材料を使うと反応後の酵素回収がしやすくなるため、連続処理や再使用を意識した研究に向いています。ただし、Enzymes.bioで提供する本品をこれらの固定化酵素として表現するものではありません。ここで重要なのは、グルコアミラーゼの性能が酵素分子そのものだけでなく、周囲の物理化学環境に左右されるという点です[10][11]。
グルコアミラーゼはタンパク質であるため、わずかな構造変化が活性や安定性に影響します。Aspergillus niger由来グルコアミラーゼの脱糖鎖化研究では、糖鎖が構造、活性、安定性に関与することが扱われています。これは、酵素を単なる化学触媒のように扱うのではなく、立体構造を保つ生体高分子として扱う必要があることを示します[3]。
別の研究では、Aspergillus awamori由来グルコアミラーゼに追加のチオール基を導入し、熱安定性と触媒活性への影響が検討されています。チオール基やジスルフィド結合はタンパク質の立体構造安定化に関係するため、こうした分子改変研究は、糖化工程で重要となる耐熱性や長時間反応性を改善するための基礎になります[12]。
近年は、機械学習による構造安定化や固定化を組み合わせて、グルコアミラーゼの活性と安定性を高める研究も行われています。これは、産業酵素が経験的な改良だけでなく、配列、構造、反応環境を統合的に設計する段階へ進んでいることを示します。ただし、こうした先端研究の成果は個別酵素や研究条件に依存するため、一般の糖化工程では、まず原料処理と工程条件の整合性を重視するのが実務的です[13]。
| 用途領域 | グルコアミラーゼの主な貢献 | 工程上の制約・注意点 |
|---|---|---|
| 発酵用糖液 | デキストリンをグルコースへ進め、発酵可能糖を増やす | 発酵微生物の条件と酵素反応条件の整合が必要 |
| バイオエタノール | でんぷん質原料から発酵基質を供給 | 液化不足や分岐残存が糖化の律速になり得る |
| 醸造・発酵食品 | アルコール生成や発酵進行を支える糖を供給 | 風味設計上、糖化を進めすぎない判断もあり得る |
| グルコースシロップ | グルコース比率の高い糖化液づくりに寄与 | 分岐デキストリンには脱分岐酵素の補助が有効な場合がある |
| 食品・飲料工程 | 糖組成、粘度、発酵性の再現性を支える | 品質・安全管理の流れに酵素管理を組み込む必要がある[5] |
この表の要点は、グルコアミラーゼの価値が「糖を増やす」だけではないことです。発酵工程では糖の供給速度、食品工程では糖組成と粘度、糖液製造では最終的なグルコース比率が重要になります。グルコアミラーゼはこれらに関与しますが、最終結果は液化、脱分岐、発酵、熱履歴、原料品質と組み合わさって決まります[1]。
Enzymes.bioは、グルコアミラーゼをB2B向けに供給するオンライン販売業者です。製造業者や研究所として酵素を開発・評価する立場ではなく、実務ユーザーが必要な酵素を入手できるよう、1kg単位で直接購入できる形態を採っています。注文時にはCoAとSDSが併せて提供されるため、受入記録、安全管理、社内保管文書として利用できます。
本品の説明では、過度な性能保証や、特定工程での収率断定を避けることが重要です。グルコアミラーゼは、でんぷん糖化と発酵基質づくりに有用な酵素ですが、実際の結果は原料、前処理、工程条件、併用酵素、発酵微生物に依存します。研究文献でも、酵素の安定性、固定化、構造改変、複合酵素化が継続的に検討されており、産業糖化は単一酵素の添加だけで完結するものではなく、工程全体の設計で最適化される分野です[6][12]。
グルコアミラーゼは、でんぷん糖化において、液化後のデキストリンやオリゴ糖をグルコースへ進める中核的な糖化酵素です。発酵用途では、微生物が利用しやすい糖を供給し、バイオエタノール、醸造、発酵食品、糖化液製造で重要な役割を果たします。α-アミラーゼによる液化、グルコアミラーゼによる末端糖化、プルラナーゼによる脱分岐の組み合わせは、でんぷん構造の複雑さに対応する基本的な考え方です[1]。
一方で、グルコアミラーゼは万能な単独処理剤ではありません。生でんぷんへの直接作用、分岐デキストリンの完全処理、高温長時間での安定性、非発酵性糖の抑制は、酵素特性と工程条件に左右されます。固定化、MOF、ナノフラワー、磁性CLEA、タンパク質改変などの研究は、こうした課題に対する科学的アプローチを示していますが、実務ではまず液化状態、糖化条件、発酵条件の整合性を取ることが重要です[8][10]。
Enzymes.bioのグルコアミラーゼは、でんぷん糖化および発酵基質づくりに利用できる糖化酵素として、1kg単位でオンライン購入できます。CoAとSDSは注文時に提供されるため、食品・飲料、発酵、でんぷん糖関連の工程管理文書に組み込めます。科学的に最も適切な理解は、「液化後のでんぷん由来デキストリンをグルコースへ進め、発酵可能糖の供給を支えるグルコアミラーゼ系糖化酵素」です。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Glucoamylase 200,000 U/G Starch Saccharification Fermentation Saccharification Enzymeを購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。