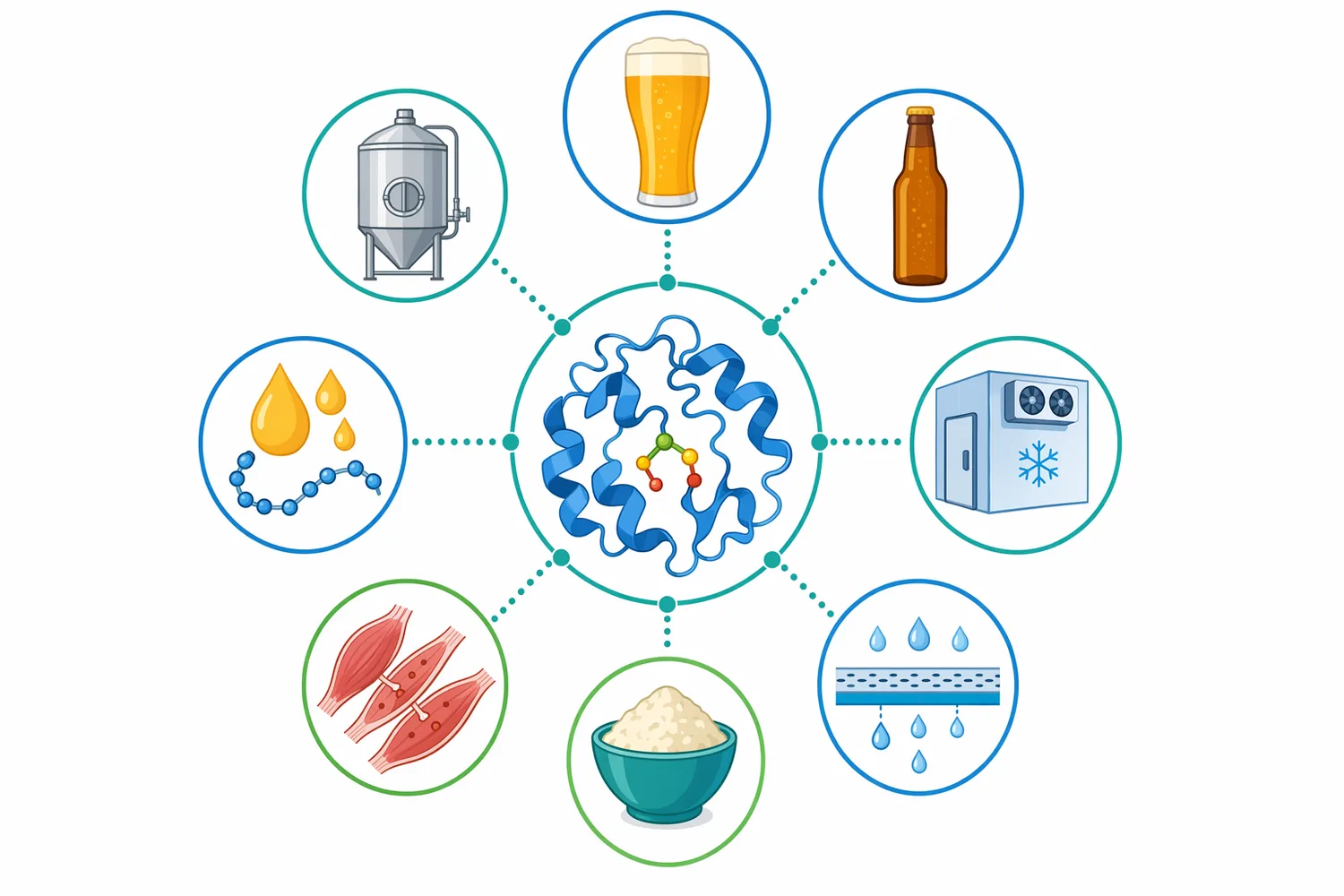
液状パパイン酵素は、ビール中に残る麦芽由来タンパク質を小さなペプチドへ加水分解し、タンパク質–ポリフェノール複合体によるチルヘイズや保存中の濁りを抑える目的で使われる清澄化補助酵素です。パパインは植物由来のシステインプロテアーゼで、ビールの外観安定性を高める一方、過度に作用すると泡安定性に関与するタンパク質にも影響し得るため、発酵後からろ過・包装前後の工程設計と組み合わせて理解する必要があります[1]。
Enzymes.bio は本製品を製造業者や研究機関としてではなく、ビール清澄化およびタンパク質加水分解用途の酵素原料を供給するオンライン販売事業者として提供しています。製品は1kg単位で直接購入でき、注文時には記録管理と安全管理に用いる CoA と SDS が併せて提供されます。
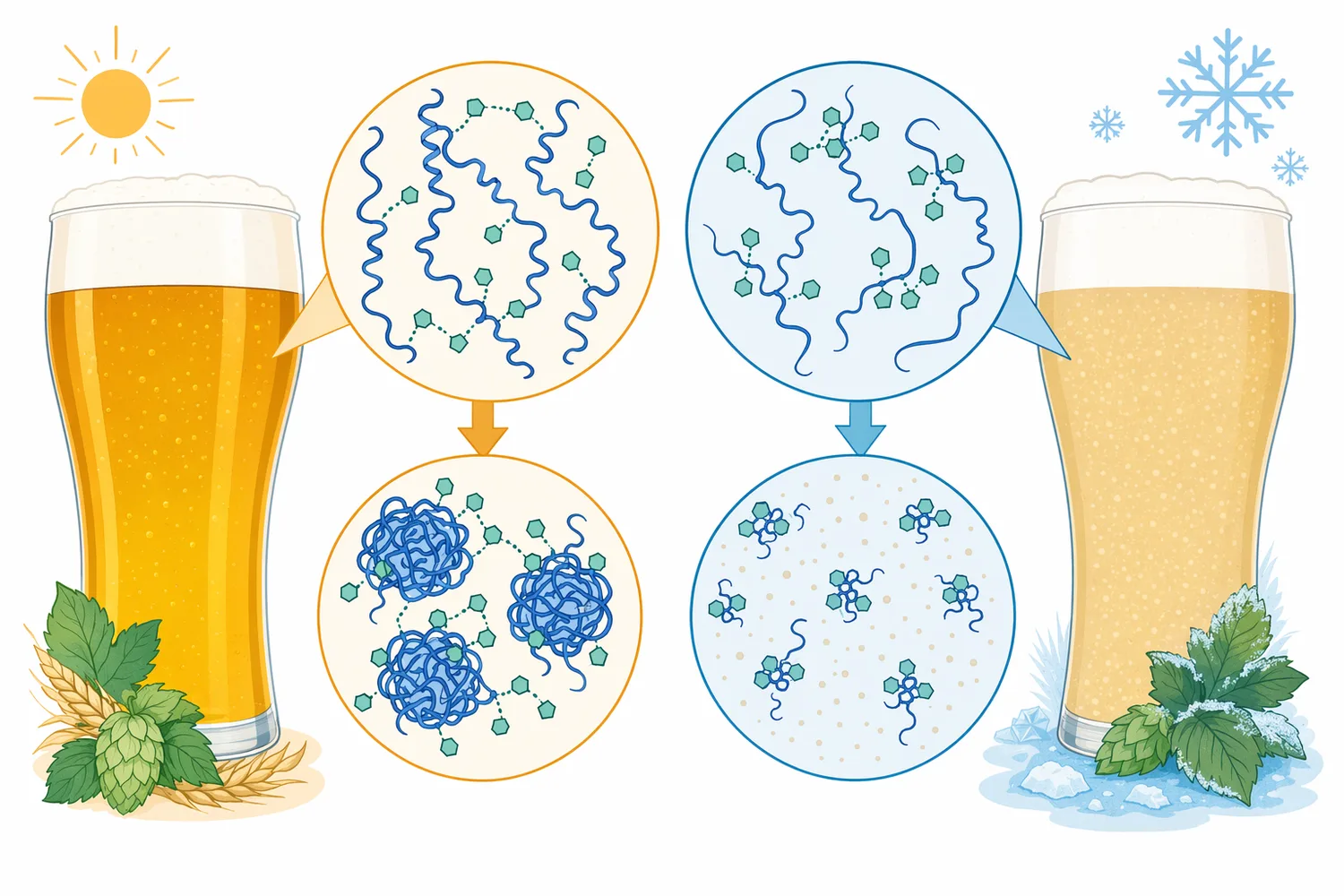
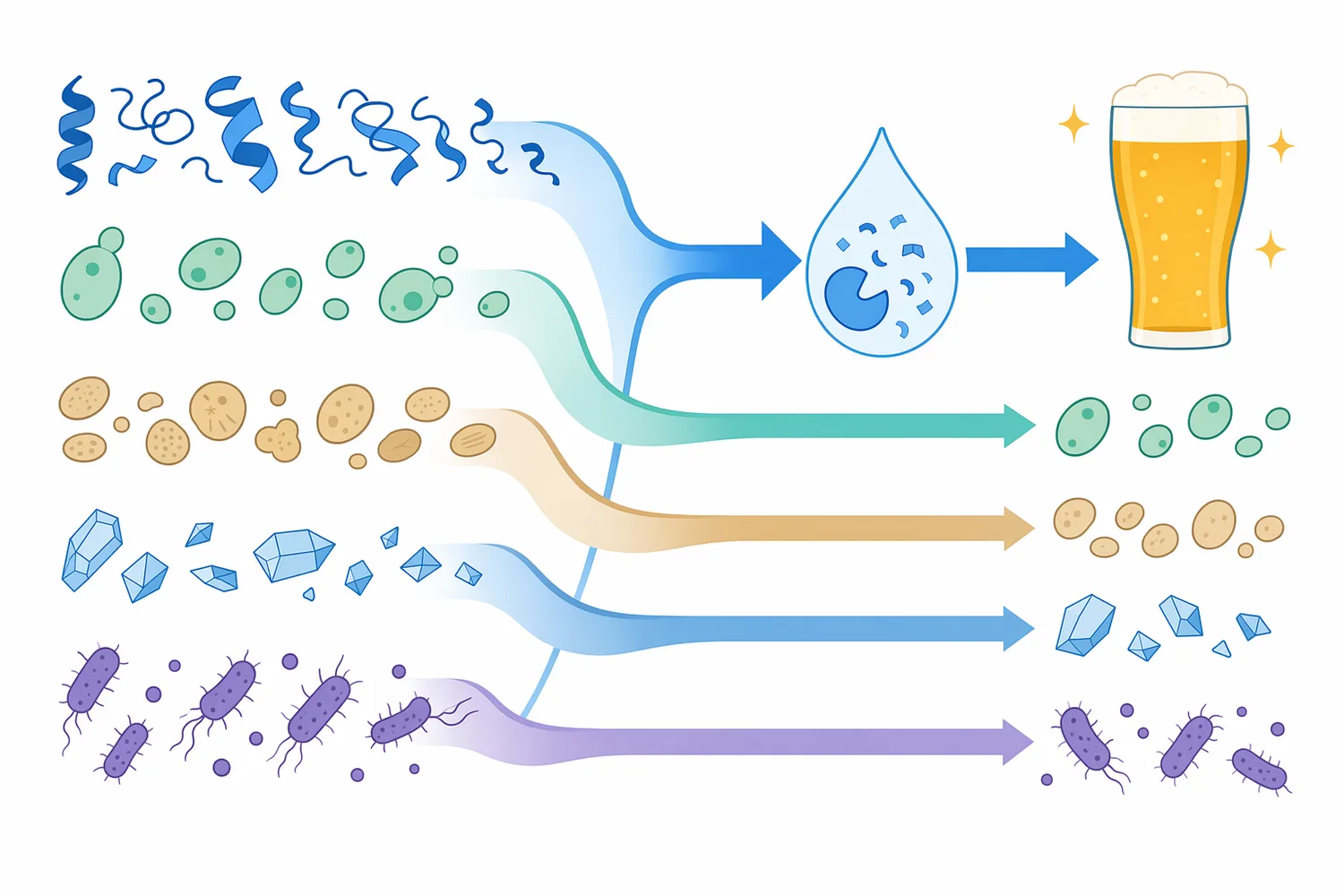
ビールの濁りは、単に「ろ過が不十分」という問題ではなく、原料タンパク質、ポリフェノール、発酵副生成物、冷却条件、保管時間が重なって起こるコロイド安定性の問題です。麦芽由来タンパク質の一部は発酵後もビール中に残り、冷却時にポリフェノールと可逆的または不可逆的な複合体を形成して、目視できる白濁や鈍い光沢低下を引き起こします。パパインは、この複合体形成に関わるタンパク質側を酵素的に小さくすることで、ビールの清澄性と外観安定性を支援するプロテアーゼです[1]。
チルヘイズは、冷えたときに目立ち、温度が上がると一部が消える初期段階から、保存中に常温でも消えにくい恒久的な濁りへ進行することがあります。タンパク質が一定以上の分子サイズと反応性を保っていると、ポリフェノールとの架橋により散乱粒子が成長しやすくなります。パパイン処理では、タンパク質鎖を切断して溶解性や相互作用性を変化させるため、物理的なろ過だけでは扱いにくい潜在的なヘイズ前駆体に対して作用できます[2]。
ビール用途でのパパインは、古典的なチルプルーフィング酵素として研究されてきました。Kennedy らの比較研究では、パパイン、キモトリプシンおよび関連タンパク質のビール冷却濁り防止能が比較され、酵素の種類によってヘイズ制御能力やビール品質への影響が異なることが示されています[1]。この点は、パパインが「タンパク質を分解すればよい」という単純な添加物ではなく、清澄化、泡、香味、貯蔵安定性のバランスに関わる工程因子であることを示しています。
パパインは、パパイヤに由来する植物性プロテアーゼとして知られ、タンパク質のペプチド結合を加水分解する酵素です。植物にはパパイン様システインプロテアーゼのファミリーが広く存在し、Arabidopsis の木部ペプチダーゼ XCP1 もパパイン様酵素との関連が検討されています[3]。このファミリーに共通する特徴は、活性中心のシステイン残基を反応の起点として用い、タンパク質鎖の特定部位を水の関与により切断する点にあります。
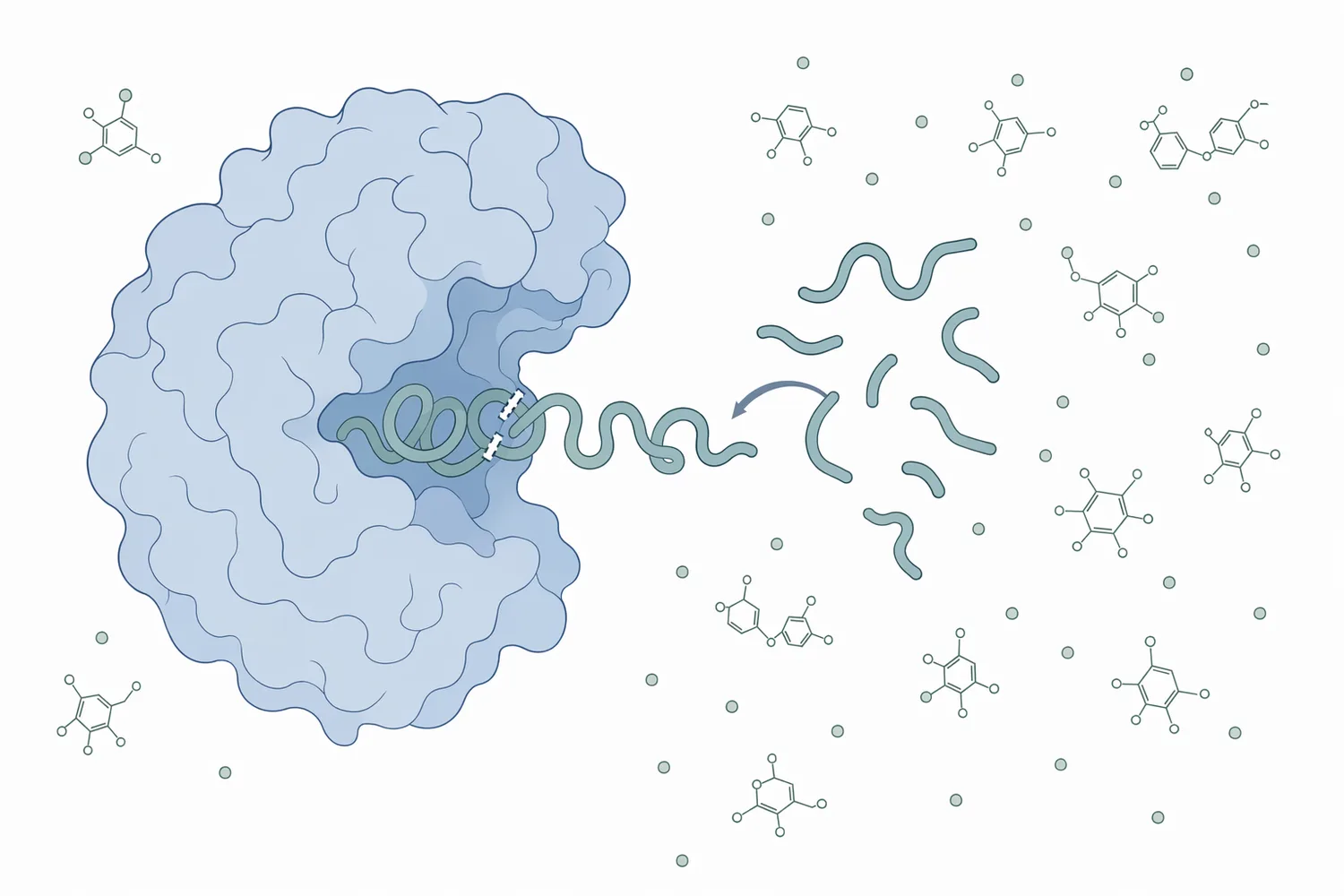
酵素反応として見ると、パパインは基質タンパク質を結合部位に保持し、活性中心でペプチド結合を切断します。一般にシステインプロテアーゼでは、求核性を持つシステイン残基がペプチド結合のカルボニル炭素へ反応し、一時的なアシル酵素中間体を経て加水分解が進みます。パパインの野生型酵素と改変体を扱った分子動力学研究は、この酵素が構造的に精密な活性部位を持つタンパク質であり、触媒機能が立体配置に強く依存することを示す関連研究として位置づけられます[4]。
この機序がビール清澄化で重要なのは、パパインがタンパク質を「沈める」のではなく、分子そのものを切断して性質を変えるからです。吸着剤がポリフェノールやタンパク質を表面へ捕捉するのに対し、パパインはタンパク質鎖の長さ、親水性、立体構造、ポリフェノールとの結合能を変化させます。そのため、同じ濁り低減という結果を目指していても、ベントナイト、シリカゲル、PVPP、タンナーゼ、プロリン特異的エンドプロテアーゼとは作用点が異なります[5]。
麦芽タンパク質は、糖化・煮沸・発酵・熟成の各工程で変性、沈殿、分解を受けますが、すべてが除去されるわけではありません。残存する中分子量タンパク質やポリペプチドには、泡を支えるもの、口当たりに寄与するもの、ヘイズ形成に関わるものが混在します。パパインはこれらを完全に選別して切断する酵素ではないため、清澄化効果と泡品質のあいだに技術的な緊張関係が生じます[6]。
ヘイズ形成に関わるタンパク質では、プロリンに富む領域や疎水性領域がポリフェノールとの相互作用に関与することがあります。プロリン特異的エンドプロテアーゼがビールのコロイド安定性とグルテン低減に利用されるのは、こうした難分解性またはヘイズ活性の高い配列に狙いを置くためです[7]。一方、パパインはより広い基質認識を示すプロテアーゼであり、特定のプロリン隣接結合だけに限定されず、幅広いタンパク質断片化を通じてヘイズ前駆体を減らす方向に働きます。
酵素処理後のタンパク質断片は、もとのタンパク質より小さなペプチドまたは遊離アミノ酸に近い形へ移行します。食品タンパク質の加水分解研究では、パパイン処理によりタンパク質の分解度、溶解性、乳化性、抗酸化性などが変化することが報告されており、酵素処理が単なる分子量低下だけでなく、表面性状や機能性の変化を伴うことが示されています[8]。ビールでは、この「分子サイズと相互作用性の変化」が濁り形成の抑制に結びつきます。
ビール清澄化では、単一の手段ですべての濁り要因を処理するより、タンパク質側、ポリフェノール側、酵母・微粒子側を工程全体で管理する考え方が現実的です。パパインはタンパク質加水分解に強みを持ちますが、ポリフェノールを直接分解する酵素ではなく、酵母細胞を除去するろ過材でもありません。果汁・飲料分野では、タンナーゼがタンニン除去を通じてヘイズ低減と品質改善に寄与することが報告されており、対象成分により使う技術が変わることが分かります[5]。
| アプローチ | 主な作用対象 | 機序の要点 | ビール品質で注目する点 |
|---|---|---|---|
| パパイン | 麦芽由来タンパク質、ポリペプチド | ペプチド結合を加水分解し、タンパク質の分子サイズと相互作用性を変える | 清澄性向上と泡安定性の両立 |
| プロリン特異的エンドプロテアーゼ | プロリンを含むヘイズ活性タンパク質、グルテン関連配列 | 特定配列への作用によりコロイド安定性やグルテン低減を狙う | ヘイズ活性タンパク質と泡活性タンパク質への選択性 |
| タンナーゼ | タンニン、ポリフェノール関連成分 | タンニンを分解・変換し、ポリフェノール側のヘイズ要因を下げる | 渋味、色調、ポリフェノールバランス |
| 物理的ろ過・遠心 | 酵母、凝集粒子、沈殿物 | 粒子サイズや密度差を利用して除去する | 透明度、歩留まり、香味保持 |
| 吸着安定化材 | タンパク質またはポリフェノール | 表面吸着によりヘイズ前駆体を減らす | 過剰除去によるボディ・香味変化 |
プロリン特異的エンドプロテアーゼについては、麦芽由来のヘイズ活性タンパク質と泡活性タンパク質への影響を調べた研究があり、清澄化酵素が泡に無関係ではないことを明確に示しています[6]。パパインでも同様に、濁りに関わるタンパク質を十分に分解しつつ、泡形成・泡保持に寄与するタンパク質を過度に失わない工程上のバランスが重要です。
ビール中のタンパク質負荷は、麦芽の種類、副原料、糖化設計、煮沸条件、発酵後処理により大きく変動します。たとえば、麦芽化していない穀物を用いる醸造では、酵素供給、可溶化、ろ過性、窒素成分の挙動が標準的な麦芽主体の設計と異なります。未麦芽大麦、麦芽、米、酵素製剤を組み合わせた小規模糖化の比較研究では、原料比率が糖化性能や麦汁特性に影響することが示されています[9]。
オーツ麦を用いた醸造でも、麦芽化オーツと未麦芽オーツでは麦汁やビール特性が変わります。オーツはタンパク質、脂質、β-グルカンなどの観点から、通常の大麦麦芽とは異なるろ過性・濁り・口当たりをもたらす可能性があります[10]。こうした原料を使うビールでは、意図的な濁りを残すヘイジースタイルと、透明性を求めるラガーやクリアエールで、パパインの意味合いが大きく変わります。
透明な製品設計では、パパインは残存タンパク質による冷却濁りを抑える手段になります。一方、濁りを外観価値として扱うスタイルでは、酵素処理により望ましいボディ感や見た目が変わる可能性があります。このため、パパインは「どのビールにも常に入れる清澄剤」ではなく、目標とする透明度、泡、口当たり、保存安定性に合わせて考えるタンパク質制御ツールです[1]。
ビールの泡は、炭酸ガスだけでなく、表面に集まるタンパク質、ポリペプチド、ホップ由来成分、多糖類などによって支えられます。泡膜を安定化するタンパク質は、気液界面で広がり、粘弾性のある膜を形成します。パパインがこうした泡活性タンパク質を過度に小さくすると、泡立ちはあっても泡持ちが短くなったり、泡のきめが粗くなったりする可能性があります[6]。
この問題は、清澄化酵素全般に共通する「選択性」の課題です。Cramer らのプロリン特異的エンドプロテアーゼ研究では、コロイド安定性改善とグルテン低減が扱われていますが、同時にビール中タンパク質のどの部分へ作用するかが重要になります[7]。パパインは広範なプロテアーゼであるため、処理目的をチルヘイズ低減に置く場合でも、泡に寄与するタンパク質を完全に無視することはできません。
実務的には、パパインの価値は「タンパク質を最大限分解すること」ではなく、「最終ビールで問題になるヘイズ活性タンパク質を、必要な範囲で減らすこと」にあります。濁りが減っても、泡、ボディ、香味が損なわれれば製品品質としては不十分です。したがって、パパインは透明性を改善する強力な選択肢であると同時に、ビールの官能品質全体の中で位置づけるべき酵素です[1]。
パパインは、ビール中に残ったタンパク質へ作用させるため、主に発酵後からろ過・包装前後の清澄化工程で位置づけられます。発酵前の麦汁段階では、糖化酵素や煮沸によるタンパク質凝固など、別のタンパク質制御がすでに起こっています。発酵後に用いる意義は、実際に最終ビールへ持ち越される可溶性タンパク質やポリペプチドを対象にできる点です[2]。
ただし、酵素は条件が合えば反応を続けるため、処理後の残存活性には注意が必要です。包装後もタンパク質分解が進むと、保存中に泡安定性、口当たり、コロイド状態が変化する可能性があります。Kennedy らの水不溶性パパイン複合体の研究は、パパインを担体へ結合させてチルプルーフィングへ使う考え方を扱っており、酵素作用を清澄化に利用しつつ、工程内での制御性を高めようとする研究的背景を示しています[2]。
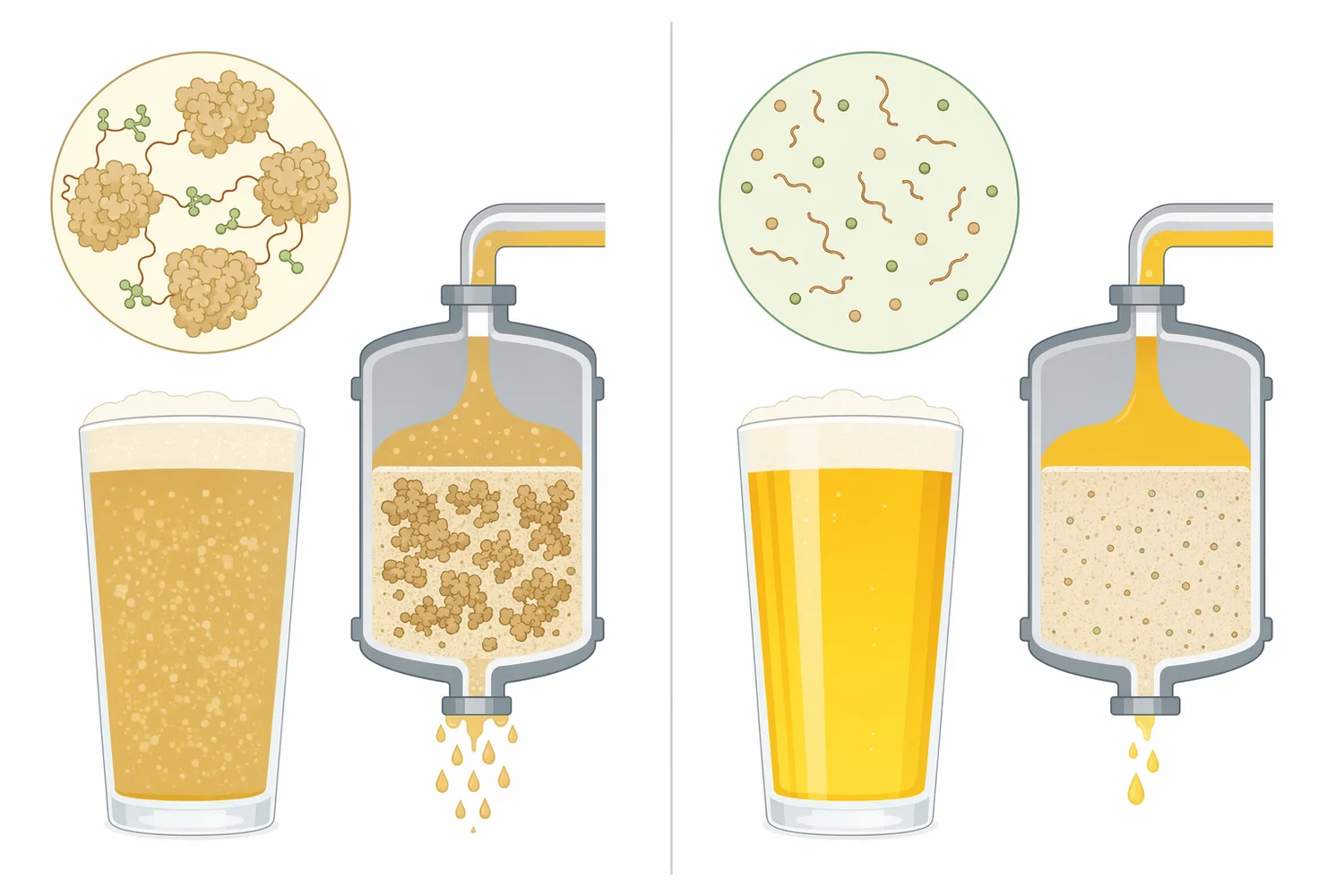
発酵後のビールは、低温、アルコール存在下、低めのpH、ホップ成分の共存という食品加工としては複雑な環境です。低温適応酵素のバイオテクノロジー研究が示すように、温度条件は酵素反応速度と基質選択性に大きく影響します[11]。パパインの使用を考える場合も、単に添加の有無ではなく、ビールの温度履歴、接触時間、後段のろ過・安定化処理と一体で捉える必要があります。
パパインはビール清澄化だけでなく、食品タンパク質をペプチドへ変換する加水分解酵素としても広く研究されています。チョウザメタンパク質の加水分解研究では、パパイン処理条件が加水分解度と機能特性に影響することが報告されており、酵素によるタンパク質改質が溶解性や食品機能を左右することが示されています[8]。この知見は、ビール中のタンパク質でも「切断の程度」が最終品質に関わることを理解する助けになります。
米タンパク質では、酵素加水分解を併用したフィブリル化処理により、溶解性、乳化性、抗酸化性が変化することが報告されています[12]。大豆タンパク質や緑豆タンパク質でも、用いるプロテアーゼの種類により、限定加水分解物の構造、界面挙動、泡特性が変わることが示されています[13]。これらはビールそのものの研究ではありませんが、パパインを含むプロテアーゼがタンパク質の界面挙動や泡関連特性へ影響し得ることを補強します。
エマルション系でも、パパインはタンパク質膜を分解することで構造安定性を変えます。ピーナッツ油体エマルションの脱乳化研究では、パパインが油体表面のタンパク質構造へ作用し、エマルション破壊に関与する機序が検討されています[14]。ビールの泡膜やコロイド粒子とは系が異なりますが、「タンパク質が界面を安定化している場合、プロテアーゼ処理はその安定性を変える」という共通原理を示しています。
ビール中のグルテン関連タンパク質を低減したい場合、近年はプロリン特異的エンドプロテアーゼが注目されています。グルテンはプロリンに富む配列を含み、一般的なプロテアーゼでは分解されにくい部位が残ることがあります。Cramer らは、コロイド安定性の改善とグルテン低減を目的とした新しいプロリン特異的エンドプロテアーゼを報告しており、配列選択性の高い酵素がビール品質上の特定課題に対応し得ることを示しています[7]。
Pijning らの研究では、Aspergillus niger 由来の酸性プロリン特異的エンドプロテアーゼによるタンパク質加水分解の構造的・時間分解的機構が調べられています[15]。このような酵素は、パパインより狭い標的特性を持つ場合があり、ヘイズ活性タンパク質やグルテン関連配列へのアプローチが異なります。したがって、パパインをグルテン低減専用酵素として説明するのは適切ではなく、主な価値は広範なタンパク質加水分解による清澄化支援にあります。
パパインは、ビール中のタンパク質全体を広く改質する酵素です。プロリン特異的酵素が特定配列を狙う「配列選択型」に近いのに対し、パパインはタンパク質量やヘイズ前駆体を総体として下げる「広域プロテアーゼ」として理解した方が実務に合います。この違いを把握することで、パパインの利点と限界を過大評価せずに扱えます[6]。
パパイン処理によりタンパク質が小さくなると、冷却時に可視光を散乱する大きな複合体が形成されにくくなります。これにより、ビールの透明度、輝き、冷蔵流通時の外観安定性が改善される可能性があります。パパインのチルプルーフィング能を扱った研究では、ビール中タンパク質への酵素作用が冷却濁り防止と関連することが検討されています[1]。
ろ過性についても、タンパク質が過剰に残るとフィルターの目詰まりやろ過差圧の上昇に関与する場合があります。パパインは酵母や不溶性粒子を直接除去するものではありませんが、可溶性またはコロイド状タンパク質を小さくすることで、後段の清澄化・ろ過負荷を下げる方向に働くことがあります。特に高タンパク質原料、未麦芽副原料、オーツや小麦を含む配合では、タンパク質挙動が透明性とろ過性の両方に関係します[10]。
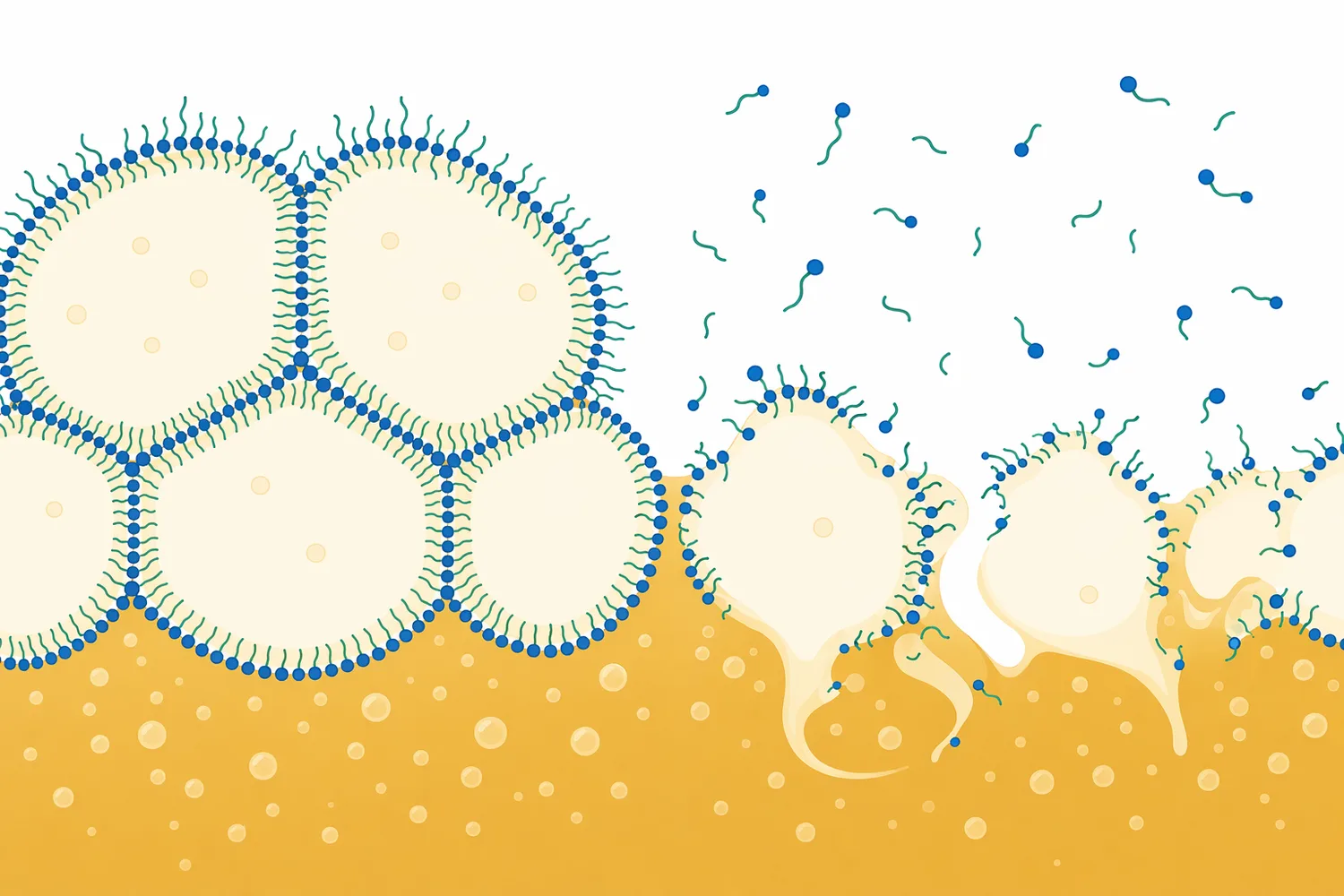
ただし、すべての濁りがパパインで解決するわけではありません。酵母残存、デンプン濁り、β-グルカン、微生物汚染、金属イオン、ポリフェノール過多など、原因がタンパク質以外に偏っている場合、パパインの効果は限定的です。タンナーゼ研究が示すように、飲料のヘイズ改善にはポリフェノール側へのアプローチが有効な場合もあり、濁りの成因に応じて技術の役割を分ける必要があります[5]。
パパインをビール清澄化に使う場合、最も重要なのは、透明性を上げるためのタンパク質分解と、泡や口当たりを守るためのタンパク質保持のバランスです。泡活性タンパク質は、ビールの見た目だけでなく、飲用時の香り立ちや飲みごたえにも関与します。プロテアーゼ種によってタンパク質構造、界面挙動、泡特性が変わることは食品タンパク質研究でも示されており、ビールでも同様の視点が必要です[13]。
もう一つの制御点は、反応をどこで止めるか、または後続工程でどの程度作用を抑えるかです。酵素は基質が残り、条件が許す限り反応を続ける可能性があります。水不溶性パパイン複合体の研究が行われた背景には、チルプルーフィング効果を得ながら、酵素を工程中で扱いやすくするという課題意識があります[2]。液状パパインを用いる場合も、既存のろ過、安定化、保管、包装条件との整合が品質を左右します。
香味面では、タンパク質分解により生じるペプチドやアミノ酸が、味の厚み、後味、熟成中の反応に影響する可能性があります。食品タンパク質加水分解物では、生成ペプチドがうま味、苦味、抗酸化性などを示すことがあり、カニ由来ペプチドの探索研究でも、タンパク質分解と呈味ペプチド生成の関係が扱われています[16]。ビールでは望ましい変化と望ましくない変化の両方があり得るため、酵素処理は外観だけでなく風味設計の一部として考える必要があります。
Enzymes.bio が扱う液状パパイン酵素は、ビール清澄化およびタンパク質加水分解用途の原料として、1kg単位でオンライン直接販売されます。Enzymes.bio は製造業者や試験機関ではなく、既存の飲料・食品加工工程で利用される酵素原料を供給する立場です。注文時には CoA と SDS が併せて提供されるため、受け入れ記録、安全管理、社内文書保管に必要な基本情報を製品と同時に扱えます。
本製品の名称にはビール清澄化、加水分解タンパク質、生物酵素といった用途語が含まれますが、実際の価値は「液状で扱える植物性プロテアーゼとして、発酵後ビール中のタンパク質を制御できること」にあります。製品仕様の詳細は注文時に提供される文書で確認する前提とし、ここでは研究文献に基づく機序、用途、品質上の考え方に焦点を置いています。
液状パパイン酵素は、ビール中の麦芽由来タンパク質を加水分解し、チルヘイズや保存中の濁りに関わるタンパク質–ポリフェノール複合体の形成を抑えるための清澄化補助酵素です。パパイン、キモトリプシンおよび関連タンパク質のチルプルーフィング比較研究や、水不溶性パパイン複合体の研究は、パパインがビールの冷却濁り防止に関連する酵素として長く検討されてきたことを示しています[1]。
一方で、パパインはヘイズ活性タンパク質だけを完全に選択して切断する酵素ではありません。泡安定性に寄与するタンパク質、ボディに関わるポリペプチド、香味に影響し得るペプチド生成も同時に考える必要があります。プロリン特異的エンドプロテアーゼ研究が示すように、ビール清澄化酵素では「どのタンパク質をどの程度変えるか」が品質を左右します[6]。
Enzymes.bio の液状パパイン酵素は、ビール清澄化とタンパク質加水分解を目的とするユーザーが、1kg単位でオンライン購入できる酵素原料です。CoA と SDS は注文時に併せて提供されます。パパインを適切に位置づけるなら、それは単なる濁り消しではなく、透明性、泡、ろ過性、保存安定性を同時に考えるためのタンパク質制御ツールです。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Papain 80,000 U/G Liquid Beer Clarification Additive Hydrolyzed Protein Biological Enzymeを購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。