Protein Hydrolysate Enzyme Neutral Protease CAS 9040-76-0 is used to convert intact proteins into shorter peptide-rich hydrolysates under near-neutral processing conditions. It is especially relevant when a buyer wants to improve protein solubility, modify functional behavior, support digestibility, or valorize protein-containing side streams without using strongly acidic or alkaline hydrolysis chemistry.
Enzymes.bio supplies this neutral protease product directly online by the 1 kg unit. The buyer pays online, the order is processed and shipped, and a Certificate of Analysis and Safety Data Sheet are provided with the order.
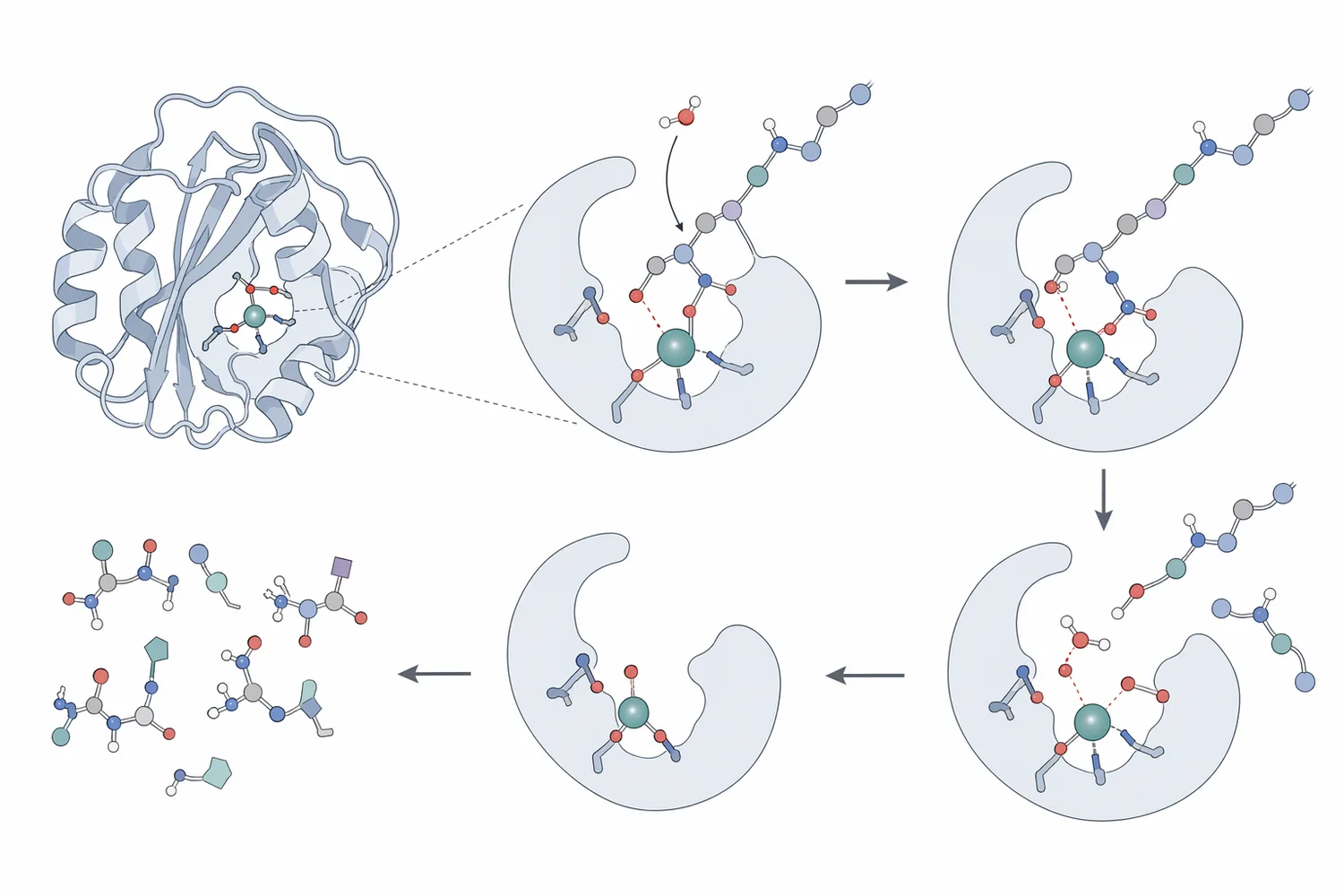
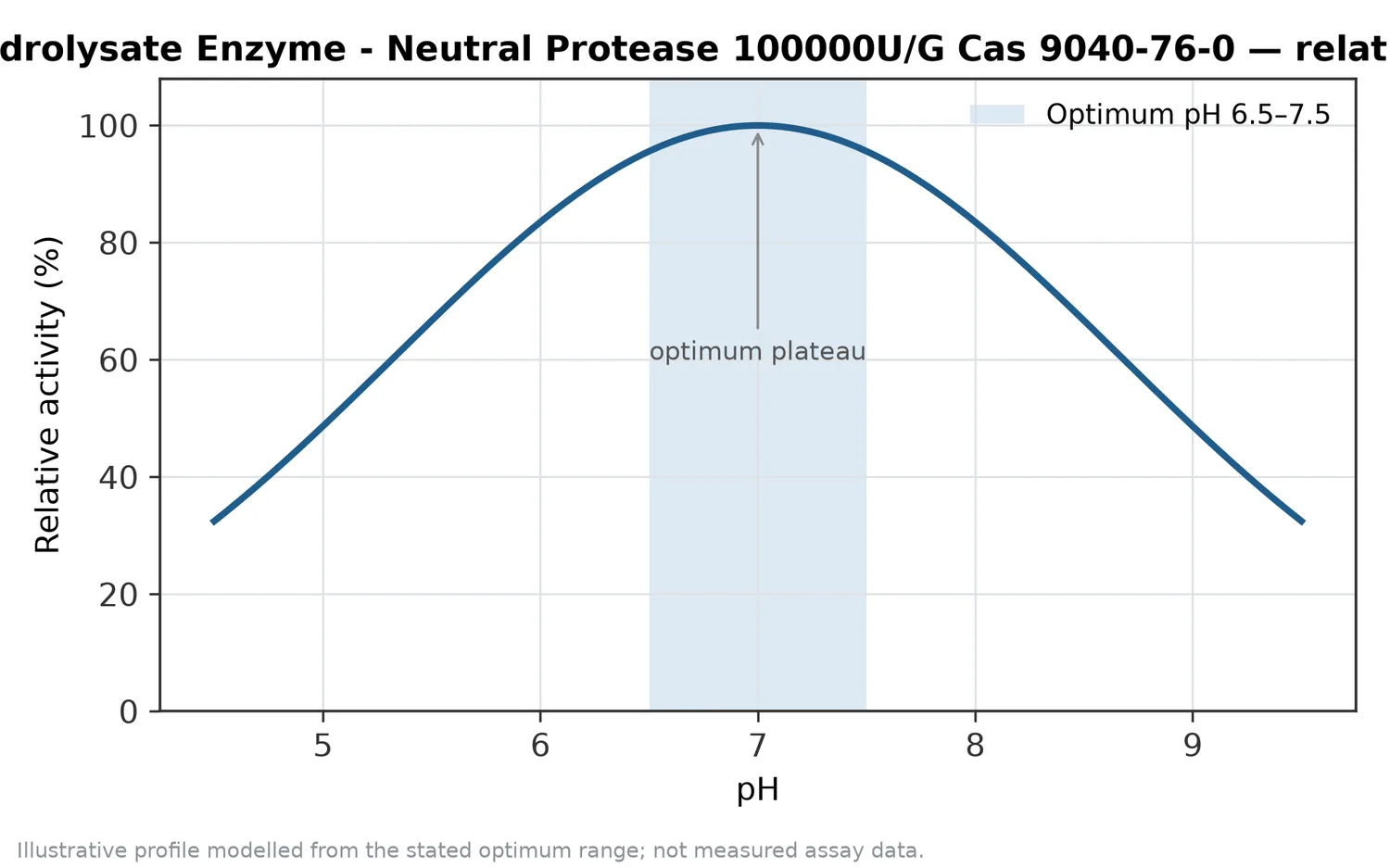
A neutral protease is a proteolytic enzyme that cleaves peptide bonds in proteins while operating best in a near-neutral environment rather than in strongly acidic or strongly alkaline systems. In protein hydrolysate production, that matters because many food, feed, fermentation, marine, dairy, plant-protein, and alternative-protein substrates are processed in water-based systems where extreme pH can damage flavor, color, nutritional quality, or downstream functionality. Broad protease reviews describe proteases as major industrial biocatalysts because they catalyze protein breakdown selectively and are used across food, detergent, leather, feed, pharmaceutical, and waste-processing applications [1].
The practical role of Protein Hydrolysate Enzyme Neutral Protease CAS 9040-76-0 is to help transform large, folded or aggregated protein molecules into mixtures of smaller peptides and amino-acid-containing fractions. This is not just a size-reduction step. When peptide bonds are cut, the protein’s three-dimensional structure loosens, buried amino-acid regions can become exposed, charged groups become more accessible to water, and the hydrolysate can behave differently in solution. These structural changes are why controlled enzymatic hydrolysis can alter solubility, viscosity, foaming, emulsification, bitterness, savory flavor development, digestibility, and the formation of bioactive peptide fractions.
Neutral protease is particularly useful when the desired outcome is controlled hydrolysis rather than aggressive destruction of the protein. Chemical hydrolysis can be harsh and less selective, while enzymatic hydrolysis can be guided by reaction conditions and substrate accessibility. Food-grade protease literature has long emphasized that microbial proteases are valuable in food biotechnology because they can modify proteins under comparatively mild conditions, supporting applications such as protein hydrolysates, flavor development, and improved processing of protein-rich materials [2].
Proteins are long amino-acid chains folded into compact structures. Some peptide bonds sit on the surface and are easy for the enzyme to access; others are hidden inside folded domains, aggregated particles, oilseed meal matrices, muscle structures, or cell-wall-rich plant materials. Neutral protease acts where the protein chain is accessible, binding to peptide segments and catalyzing bond cleavage. Each cleavage creates shorter chains with new terminal amino and carboxyl groups, which can increase hydration and change charge distribution.
As hydrolysis proceeds, several concrete changes occur in the substrate:
The importance of enzyme choice is well illustrated in oyster protein hydrolysate research, where different proteases produced hydrolysates with different molecular-weight profiles, antioxidant activity, and cellular protective effects. The key lesson for industrial use is that protease hydrolysis is not a generic “cutting” reaction; the enzyme, substrate, and process environment together shape the peptide distribution and final function [3].
Proteases are often discussed by the pH environment in which they perform best. Acid, neutral, and alkaline proteases can all hydrolyze proteins, but they tend to fit different process contexts. Neutral protease is commonly attractive when the substrate, equipment, downstream formulation, or sensory target favors a moderate processing environment rather than strongly acidic or alkaline conditions. Reviews of alkaline proteases, for example, emphasize their importance in detergent, leather, waste treatment, and other processes that tolerate or require alkaline operation, whereas food-grade protease use often depends on compatibility with the food matrix and intended processing conditions [4].
| Protease type | Conceptual operating environment | Typical substrate impact | Common processing fit | Practical caution |
|---|---|---|---|---|
| Acid protease | Acidic systems | Hydrolyzes proteins under low-pH conditions; useful where the matrix is already acidic | Acidified foods, some feed or fermentation systems, protein digestion models | Low pH can affect flavor, color, minerals, and downstream neutralization needs |
| Neutral protease | Near-neutral systems | Produces peptide-rich hydrolysates while avoiding strongly acidic or alkaline treatment | Plant proteins, dairy systems, seafood by-products, alternative proteins, savory hydrolysates, feed ingredients | Outcome depends strongly on substrate accessibility and hydrolysis control |
| Alkaline protease | Alkaline systems | Strong protein-degrading action in high-pH environments | Detergents, leather processing, some waste-treatment and industrial cleaning applications | High pH may be unsuitable for sensitive food proteins or flavor systems |
This comparison is conceptual rather than a specification sheet. In real processing, the final hydrolysate depends on the protein source, pretreatment, water content, mixing, residence time, heating history, and downstream handling. Protease reviews consistently show that industrial protease performance is application-dependent, which is why the same broad enzyme class can be used in very different sectors with different outcomes [5].
Plant proteins are one of the largest opportunity areas for neutral protease because many plant-derived proteins are abundant but functionally constrained. Soybean meal, mung bean, quinoa, rice bran, sesame meal, and other plant streams often contain proteins embedded in starch, fiber, phenolics, lipids, or insoluble protein bodies. Enzymatic hydrolysis can loosen these structures, reduce molecular size, and create peptide fractions with different solubility or stability.
Mung bean protein hydrolysate research is a good example of how hydrolysis can affect both physicochemical behavior and oxidative stability. Recent work on mung bean protein hydrolysates examined physicochemical properties and lipid-peroxidation inhibition, showing why legume hydrolysates are studied not only as protein ingredients but also as functional systems that may protect lipid-containing formulations from oxidative deterioration [6].
Rice bran is another protein-rich side stream where hydrolysis can generate higher-value fractions. Defatted pigmented rice bran protein hydrolysates have been studied for antioxidant, antidiabetic, and antihypertensive activities, reflecting the broader interest in converting cereal by-products into peptide-containing ingredients with measurable in vitro bioactivities [7]. These findings should be interpreted as substrate- and process-specific, but they support the general value proposition of enzymatic hydrolysis for underused plant protein streams.
Quinoa protein hydrolysates have also been investigated for antioxidant properties and cytoprotection against oxidative stress. This type of study is relevant because quinoa proteins are nutritionally attractive, but hydrolysis can further change their peptide profile and biological activity in model systems [8]. For buyers working with plant proteins, the practical takeaway is that neutral protease can be part of a process strategy for improving dispersibility, creating smaller peptide fractions, and exploring functional hydrolysate properties.
Soybean meal fermentation provides another useful industrial reference point. A 2025 study on Bacillus amyloliquefaciens LX-6 focused on improving neutral protease activity and applying the strain in soybean meal fermentation, connecting neutral protease production directly with soybean meal upgrading [9]. Fermentation and proteolysis can work together: microbial growth modifies the matrix, while proteases break proteins into smaller peptides that can influence nutritional value, palatability, and process performance.
Alternative proteins often need functional modification before they perform well in finished products. Microalgae, insect pupae, and other emerging protein sources can contain valuable amino-acid profiles, pigments, lipids, polysaccharides, chitin, or cell-wall structures that complicate extraction and formulation. Neutral protease can help unlock protein value by generating soluble peptide fractions and changing the way the material hydrates or disperses.
Microalgae protein hydrolysates prepared by neutral protease digestion have been studied for antioxidant properties, demonstrating direct use of neutral protease in converting algal proteins into peptide-rich hydrolysates with measurable radical-scavenging or oxidative-stability-related behavior in experimental systems [10]. For ingredient developers, this supports the idea that neutral protease is not limited to conventional soy or dairy substrates; it can also be relevant for newer biomass streams.
Insect proteins are also increasingly studied as sustainable protein sources. Silkworm pupae protein treated with a novel neutral protease has been investigated for protein hydrolysate production, showing how neutral protease can be applied to insect-derived protein material [11]. The mechanism is the same as in other protein systems—peptide bond cleavage—but the matrix differs: insect biomass may include chitinous material, lipids, pigments, and heat-affected proteins, all of which can influence enzyme accessibility and final hydrolysate properties.
Marine proteins are attractive for hydrolysate production because fish, shellfish, and sea cucumber processing can generate protein-rich residues that are difficult to use in their intact form. Controlled proteolysis can convert these residues into soluble peptides for flavor systems, nutritional ingredients, feed ingredients, pet food palatants, or functional food research. In oyster protein hydrolysate work, protease selection influenced molecular weight, antioxidant activity, and cellular protective effects, demonstrating how strongly enzyme choice can shape the final peptide profile [3].
Sea cucumber intestinal protein hydrolysates provide a more specialized example. Research has screened and characterized antifreeze peptides from sea cucumber intestinal protein hydrolysates, showing that enzymatic hydrolysis can release peptide sequences with ice-crystal-related functionality in model systems [12]. This is a clear mechanism-driven application: the hydrolysate is not valuable merely because it is “broken protein,” but because particular peptide fractions can interact with ice growth and recrystallization behavior.
A related study isolated different characterized fractions from sea cucumber intestinal protein hydrolysates and evaluated cryoprotective activity against salmon. That work connects peptide hydrolysates with frozen seafood quality, where small peptides may help protect muscle proteins from freeze-related denaturation and texture loss [13]. These findings are highly application-specific, but they illustrate the breadth of protein hydrolysate functionality that can emerge from controlled proteolysis.
In dairy and fermented foods, proteolysis is central to texture and flavor development. Caseins and whey proteins can be converted into peptides and amino-acid-containing fractions that affect mouthfeel, bitterness, umami, ripening character, and nutritional properties. A 2024 study on enzyme-modified cheddar cheese examined the molecular mechanism of cheese flavor compensation through the synergistic action of cell-free extracts and enzyme systems, showing how enzyme-driven protein breakdown contributes to flavor formation [14].
For savory hydrolysates, the mechanism is similar: neutral protease releases peptides that can contribute brothiness, body, kokumi-like fullness, or bitterness depending on peptide sequence and hydrolysis extent. Short peptides with hydrophobic amino acids can taste bitter, while amino acids and certain peptides can support savory or fermented notes. This is why controlled hydrolysis is important; more hydrolysis is not automatically better if the application requires a clean flavor profile.
One of the most common reasons to use a protein hydrolysate enzyme is to improve how a protein behaves in water. Large proteins may aggregate, sediment, form lumps, or show poor hydration. When neutral protease cuts the protein into smaller peptide fragments, those fragments can expose more ionizable and polar groups to water. The result can be improved dispersibility and reduced visible insoluble material, particularly when the original limitation was protein aggregation rather than non-protein fiber or mineral content.
This effect is not unlimited. If the raw material contains intact cell walls, high fiber, heat-damaged aggregates, or crosslinked proteins, the protease can only act on accessible peptide bonds. In plant and fermentation by-products, mechanical size reduction, hydration, heat history, and matrix structure can strongly affect how much protein the enzyme can reach. The soybean meal fermentation study involving neutral protease activity is relevant here because it shows that protein upgrading often involves both biological matrix modification and proteolytic breakdown [9].
Hydrolysate performance is closely tied to peptide size distribution. Larger peptides may retain some emulsifying or film-forming capacity, while smaller peptides may be more soluble, less viscous, and more rapidly absorbed in nutritional contexts. Excessive hydrolysis, however, can reduce structure-building ability and may increase bitterness. Research comparing proteases in oyster protein hydrolysates highlights that different enzymes generate different molecular-weight profiles, which then influence antioxidant and cellular effects [3].
Neutral protease is therefore useful when a moderate process environment and controlled peptide generation are desired. The enzyme cuts peptide bonds, but the final molecular-weight distribution is shaped by the substrate’s folding, accessibility, and reaction progression. In practice, this means hydrolysates can differ significantly even when the same broad enzyme class is used on different proteins.
Many studies evaluate protein hydrolysates for antioxidant activity because peptide sequences can donate electrons, chelate metals, scavenge radicals, or protect lipids and cells in model systems. Microalgae protein hydrolysates prepared by neutral protease digestion have been studied for antioxidant properties, supporting the link between neutral protease hydrolysis and peptide fractions with measurable antioxidant behavior [10]. Similar research exists for quinoa, rice bran, oyster, mung bean, and other protein hydrolysates.
These findings are promising but should be read carefully. Antioxidant, antidiabetic, antihypertensive, antifreeze, or cytoprotective activities are not automatic outcomes of neutral protease use. They depend on the protein’s amino-acid sequence, the peptide bonds cleaved, the final peptide mixture, concentration, purification, and the test model used. For commercial claims, finished-product testing and regulatory review are essential.
Hydrolysis can improve or weaken functional performance depending on how far the reaction proceeds. For foaming, smaller and more flexible peptides may move quickly to the air–water interface, unfold, and reduce surface tension. For emulsions, peptides need enough amphiphilic character—hydrophobic regions to interact with oil and hydrophilic regions to remain water-compatible. If hydrolysis goes too far, peptides may become too small to form strong interfacial films, reducing emulsion stability.
This is why neutral protease is often used for controlled modification rather than complete digestion. The desired endpoint may be a hydrolysate that is still functional enough to stabilize interfaces but small enough to hydrate quickly and reduce grittiness or sedimentation. Plant protein hydrolysate research, including studies on mung bean and other legumes, supports the concept that enzymatic hydrolysis changes both physicochemical behavior and oxidative stability in ways relevant to formulated foods [6].
Plant-based beverages, nutritional powders, meat alternatives, sauces, and high-protein foods often require proteins that hydrate efficiently and remain stable during processing. Neutral protease can help reduce large protein aggregates and create peptide-rich fractions that disperse more easily. This can be valuable for soy, mung bean, rice bran, quinoa, pea, and other plant-derived streams, especially when the goal is to improve use of a protein by-product or reduce processing losses.
Plant protein hydrolysates are also studied for biological activities beyond basic nutrition. Defatted pigmented rice bran protein hydrolysates have been examined for antioxidant, antidiabetic, and antihypertensive activities, while quinoa protein hydrolysates have been investigated for antioxidant properties and cytoprotection against oxidative stress [7]. These studies demonstrate the research direction, but the commercial value still depends on the intended food category, sensory profile, dosage in the finished product, and applicable claims framework.
In feed and fermentation contexts, proteolysis can make protein fractions more accessible and may improve the value of soybean meal or similar ingredients. Smaller peptides can be relevant for palatability, microbial fermentation, and nutritional utilization. The study on Bacillus amyloliquefaciens LX-6 connects improved neutral protease activity with soybean meal fermentation, supporting the use of neutral protease activity in feed-ingredient upgrading [9].
The mechanism is straightforward: large storage proteins in soybean meal are partially cleaved into peptides, while fermentation can alter antinutritional factors, texture, and matrix accessibility. A neutral protease product may be used where the process environment is closer to neutral and where the buyer wants enzymatic protein modification rather than strongly acidic treatment.
Fish, shellfish, and sea cucumber processing generate protein-rich streams that can be difficult to sell or formulate in their raw form. Neutral protease can convert such materials into soluble peptide hydrolysates that may be used in flavor bases, feed ingredients, pet food palatants, or functional hydrolysate research. Sea cucumber intestinal protein hydrolysates are a clear example of how marine by-products can be converted into peptide fractions with specialized properties such as antifreeze activity [12].
The economic appeal is that enzymatic hydrolysis can transform a variable, perishable, or low-value residue into a more uniform liquid or dried hydrolysate. The technical challenge is that marine materials may contain lipids, minerals, pigments, endogenous enzymes, and odor precursors, all of which influence final quality. Neutral protease addresses the protein fraction; overall product quality still depends on raw-material freshness and downstream handling.
Hydrolysates are not limited to nutrition or solubility applications. In frozen seafood systems, peptides may help protect muscle proteins from freeze-induced denaturation, drip loss, and textural deterioration. Sea cucumber intestinal protein hydrolysate fractions have been studied for cryoprotective activity against salmon, linking peptide composition with frozen-fish quality protection [13].
Mechanistically, cryoprotective peptides can interact with water, ice crystals, or muscle protein surfaces. They may help moderate ice recrystallization or reduce structural damage during frozen storage. This is a specialized application area and not a universal result of hydrolysis, but it shows why controlled peptide generation can create value beyond simple protein digestion.
Neutral protease can also support flavor development by releasing peptides and amino acids from dairy, meat, yeast, legume, or marine proteins. In cheese and enzyme-modified dairy systems, proteolysis contributes to ripening and flavor complexity. The enzyme-modified cheddar cheese study highlights how enzyme systems and cell-free extracts can act together to compensate and develop flavor at the molecular level [14].
In savory hydrolysates, the desired endpoint is often a balance: enough hydrolysis to release flavor-active peptides and amino acids, but not so much that bitterness dominates. Neutral conditions can be attractive when the buyer wants a gentler environment for flavor-sensitive materials.
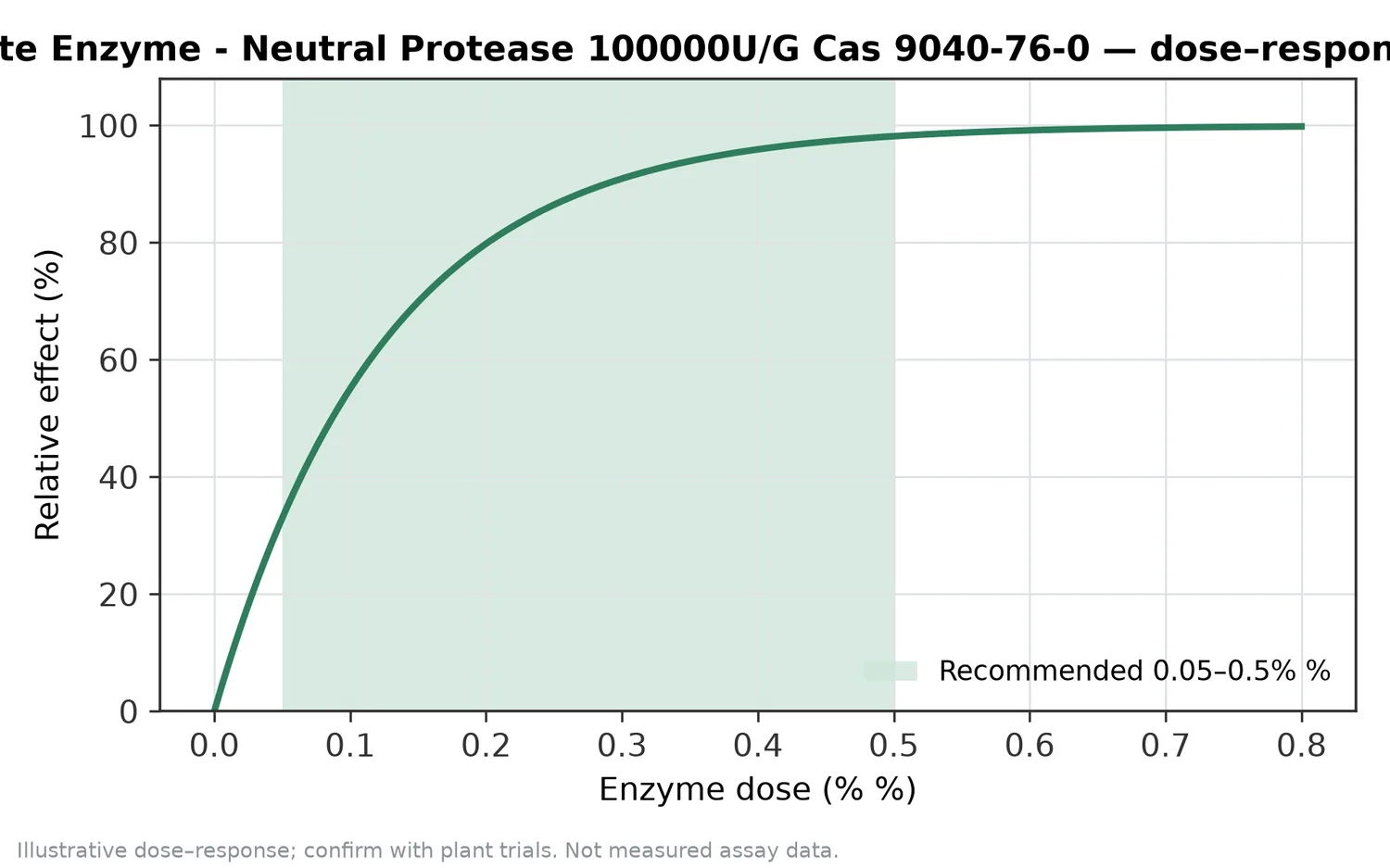
The first determinant is protein accessibility. A purified protein isolate exposes more peptide bonds than an intact seed meal, algae cell, insect meal, or fish-processing residue. If the enzyme cannot reach the protein, hydrolysis will be limited no matter how much protein is present. Milling, hydration, prior heat treatment, fermentation, and the presence of fiber or cell walls all influence accessibility.
The second determinant is hydrolysis progression. Early hydrolysis usually reduces large protein particles and can improve dispersion. Intermediate hydrolysis may create functional peptides with good solubility and interfacial behavior. Extended hydrolysis can produce smaller peptides and free amino acids, which may be desirable for digestibility or flavor intensity but may reduce structure-building properties or increase bitterness. This progression explains why protein hydrolysate production is a controlled conversion rather than a simple “add enzyme” step.
The third determinant is substrate composition. Oilseed meals, dairy proteins, seafood residues, algae, and insect proteins contain different amino-acid sequences and non-protein components. Because proteases act on peptide bonds within specific structural contexts, two substrates treated under similar conditions can produce hydrolysates with very different sensory, nutritional, and functional profiles. The broad literature on protease applications shows that substrate identity is one of the main reasons proteases are considered versatile but application-specific biocatalysts [1].
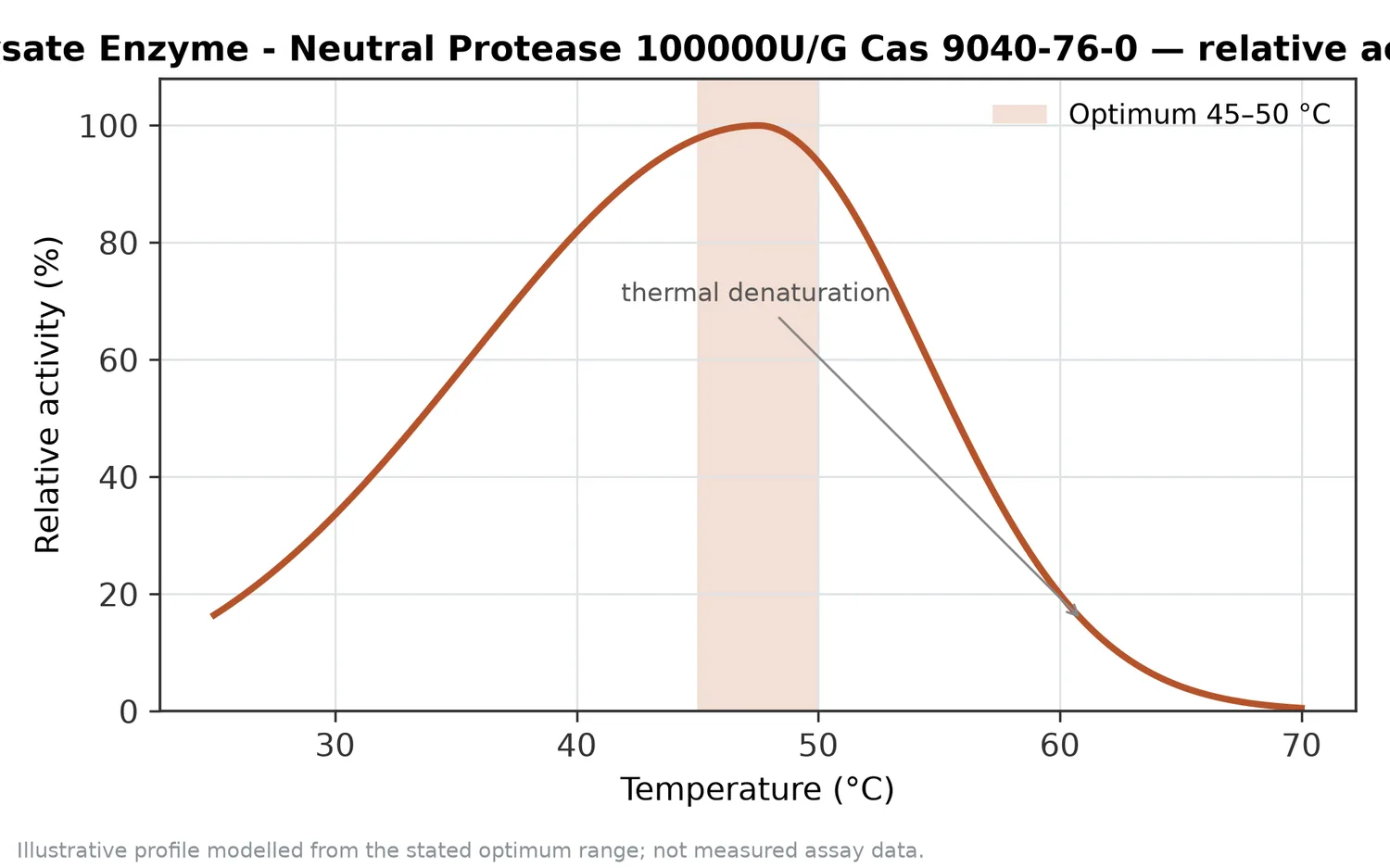
The fourth determinant is downstream handling. Heating, filtration, concentration, drying, blending, and storage can change hydrolysate appearance and performance. Peptides may participate in Maillard reactions with reducing sugars during heating, which can deepen color and create roasted or savory notes. Lipids in marine or oilseed substrates may oxidize if not controlled, which can affect odor and flavor. Neutral protease creates the peptide profile, but downstream processing determines how that profile is preserved or transformed.
Neutral protease can reasonably be expected to support protein molecular-size reduction, peptide generation, improved dispersibility in suitable substrates, and better utilization of certain protein-rich side streams. It may also support functional changes such as altered foaming, emulsification, flavor development, or oxidative-stability behavior, depending on substrate and process design. Research on microalgae, mung bean, rice bran, quinoa, oyster, silkworm pupae, soybean meal, and sea cucumber hydrolysates collectively supports the broad usefulness of protease-driven hydrolysis across diverse protein materials [10].
At the same time, neutral protease should not be treated as a universal fix for every protein-processing problem. If poor solubility is caused primarily by insoluble fiber, mineral complexes, oxidized lipids, excessive heat damage, or non-protein impurities, protease alone may not deliver the desired improvement. If the target is a bioactive claim—such as antioxidant, antihypertensive, antifreeze, antidiabetic, or cytoprotective activity—the finished hydrolysate must be evaluated in the appropriate model and regulatory context.
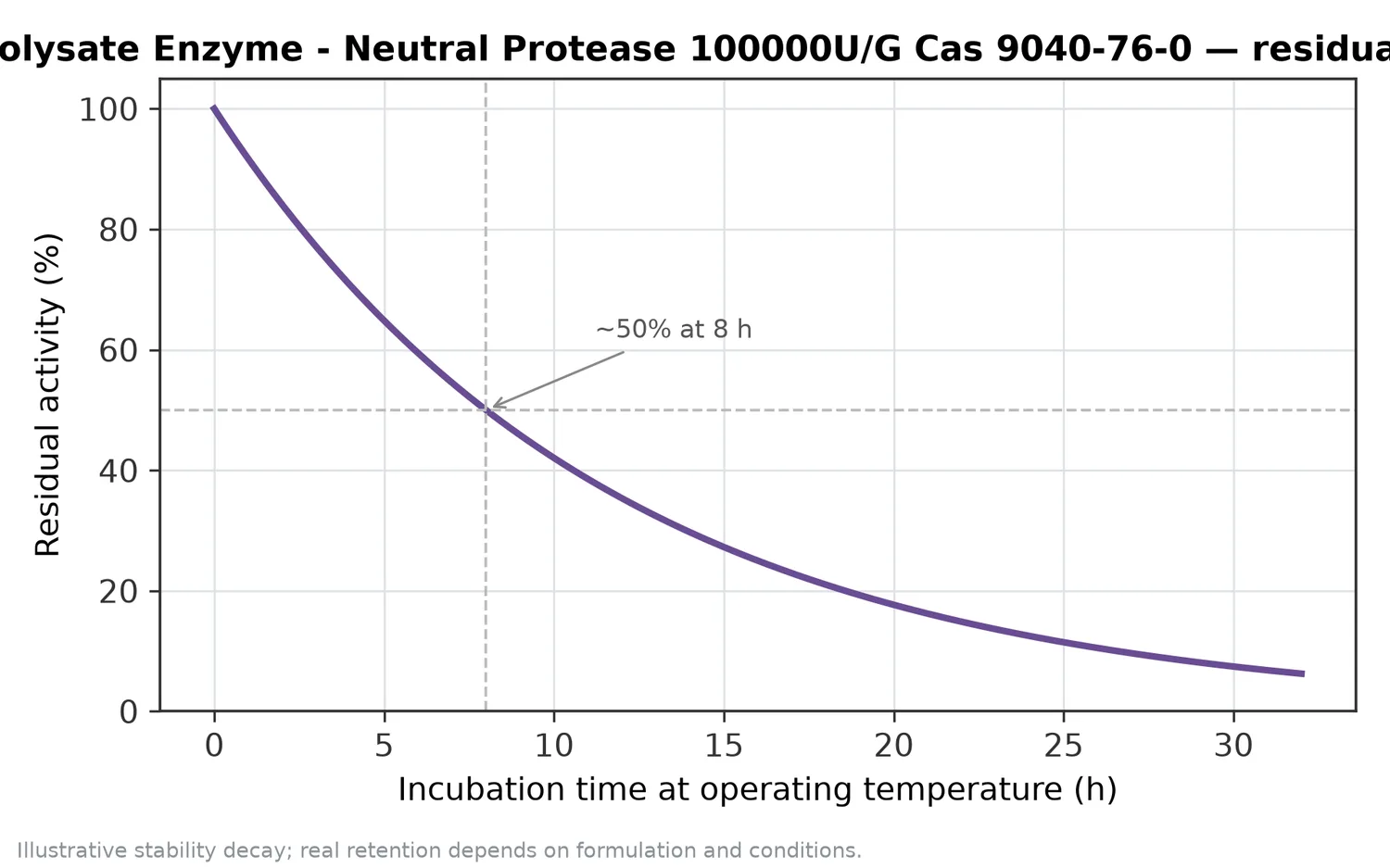
The most defensible way to view this enzyme is as a controlled protein-conversion tool. It changes proteins by cleaving peptide bonds; those cleavages change molecular size, hydration, charge exposure, peptide sequence distribution, surface behavior, and flavor potential. The value comes from aligning that conversion with the substrate and intended product format.
Enzymes.bio supplies Protein Hydrolysate Enzyme Neutral Protease CAS 9040-76-0 directly online by the 1 kg unit. The purchasing path is straightforward: the buyer places the order online, pays online, and the order is processed and shipped. A Certificate of Analysis and Safety Data Sheet are provided with the order .
Enzymes.bio is a supplier, not the manufacturer and not a testing laboratory. The product listing is intended for buyers who already know they need a neutral protease for protein hydrolysate work and want a direct online purchase route rather than a quote-based process.
Protein Hydrolysate Enzyme Neutral Protease CAS 9040-76-0 is used to produce peptide-rich hydrolysates from plant, marine, dairy, feed, fermentation, insect, algae, and other protein-containing materials under near-neutral processing conditions. Its core action is peptide-bond cleavage: large proteins are converted into smaller peptides, which can change solubility, dispersibility, interfacial behavior, flavor development, digestibility, and potential bioactivity.
The strongest evidence supports neutral protease as a versatile protein-modification enzyme. Studies on microalgae, silkworm pupae, mung bean, rice bran, quinoa, oyster, soybean meal, cheese systems, and sea cucumber hydrolysates show how protease-driven hydrolysis can create functional peptide fractions in very different substrates. Specific outcomes depend on the protein source and process, but the underlying mechanism is consistent: controlled enzymatic cutting changes the structure and performance of the protein material.
Sold by the 1 kg unit, in stock and ready to ship. Order directly on our store — pay online and we process your order. A Certificate of Analysis and Safety Data Sheet are included with every order.
Buy Protein Hydrolysate Enzyme - Neutral Protease 100000U/G Cas 9040-76-0 →Numbered in order of first citation. Open-access sources, each verified reachable at publication; citation numbers in the text link here.