Protein Hydrolysate Enzyme Neutral Protease(CAS 9040-76-0)は、中性付近の条件でタンパク質のペプチド結合を切断し、より短いペプチドや遊離アミノ酸を生成するためのタンパク質加水分解向け酵素です。植物性タンパク質、乳タンパク質、魚介・畜肉タンパク質、発酵原料などで、溶解性、分散性、呈味、発酵利用性、消化性に関わる分子サイズ分布を調整する目的で使われます。Enzymes.bioは本製品を1kg単位でオンライン販売する供給業者であり、CoAおよびSDSは注文時に併せて提供されます。
Protein Hydrolysate Enzyme Neutral Protease(CAS 9040-76-0)は、タンパク質加水分解物の製造、食品・発酵素材の前処理、植物性タンパク質や動物性タンパク質の機能性調整に使われる中性プロテアーゼ製品です。ここでいう中性プロテアーゼは、単一の生化学的ファミリー名ではなく、中性付近でタンパク質を加水分解する酵素群を実務的に指す表現です。中性プロテアーゼには、金属プロテアーゼ、セリンプロテアーゼなど異なる触媒機構を持つ酵素が含まれ得るため、名称は「どのpH領域で扱いやすいか」と「タンパク質をどのように切るか」を合わせた産業上の分類として理解するのが適切です[1]。
Enzymes.bioは製造業者や研究所ではなく、酵素をオンラインで供給するB2Bサプライヤーです。本製品は1kg単位でオンライン直接販売され、注文処理後に配送されます。試験成績書(CoA)と安全データシート(SDS)は注文時に併せて提供されますが、本稿では製造元固有の活性単位、分析法、グレード設計、単位定義には踏み込みません。この記事の焦点は、Neutral Protease(中性プロテアーゼ)がタンパク質加水分解でどのような役割を持ち、どの用途で意味を持つかを、公開文献に基づいて整理することにあります。
タンパク質原料は、栄養価が高い一方で、加工工程では水への溶けにくさ、分散不良、沈殿、粘度上昇、凝集、粉末再溶解性の低さ、発酵中の窒素源利用不足、特有の青臭さや豆臭、苦味前駆体、加熱後のざらつきなどの問題を起こします。酵素加水分解は、タンパク質の一次構造を部分的に切断し、分子量分布、表面疎水性、電荷分布、立体構造を変えることで、これらの実務課題を調整する技術です。植物タンパク質の加水分解に関するレビューでは、酵素処理が溶解性、乳化性、起泡性、ゲル化、保水・保油性などのテクノ機能性を変える手段として整理されています[2]。
中性プロテアーゼの利点は、強酸性や強アルカリ性に大きく寄せずに、比較的穏やかな条件でタンパク質の分解を進められる点です。極端なpHは、食品原料の色調、香味、ミネラル状態、他成分との相互作用、後工程の中和負荷に影響します。中性域で働くプロテアーゼは、発酵原料、乳タンパク質、豆類タンパク質、魚介タンパク質、酵母タンパク質など、pHを大きく変えたくない原料系で使いやすい選択肢になります。豆類タンパク質では、酵素加水分解により構造がほどけ、埋もれていた疎水性・親水性領域が露出し、界面挙動や水和性が変化することが報告されています[3]。
実務上は、プロテアーゼ処理の目的を「タンパク質を完全にアミノ酸へ分解すること」と考えるより、「狙った分子サイズ、呈味、溶解性、発酵適性に近づけること」と捉える方が正確です。低分子化が進むほど溶解性や発酵利用性が向上する場合がありますが、同時に疎水性ペプチド由来の苦味、過度な粘度低下、ゲル形成能の低下、乳化安定性の変化も起こり得ます。したがって、Neutral Proteaseは万能な分解剤ではなく、タンパク質素材の分子設計に使う加工酵素として位置づけるべきです。植物タンパク質加水分解のレビューでも、加水分解度やペプチド分布が機能性と用途適性を左右することが強調されています[2]。
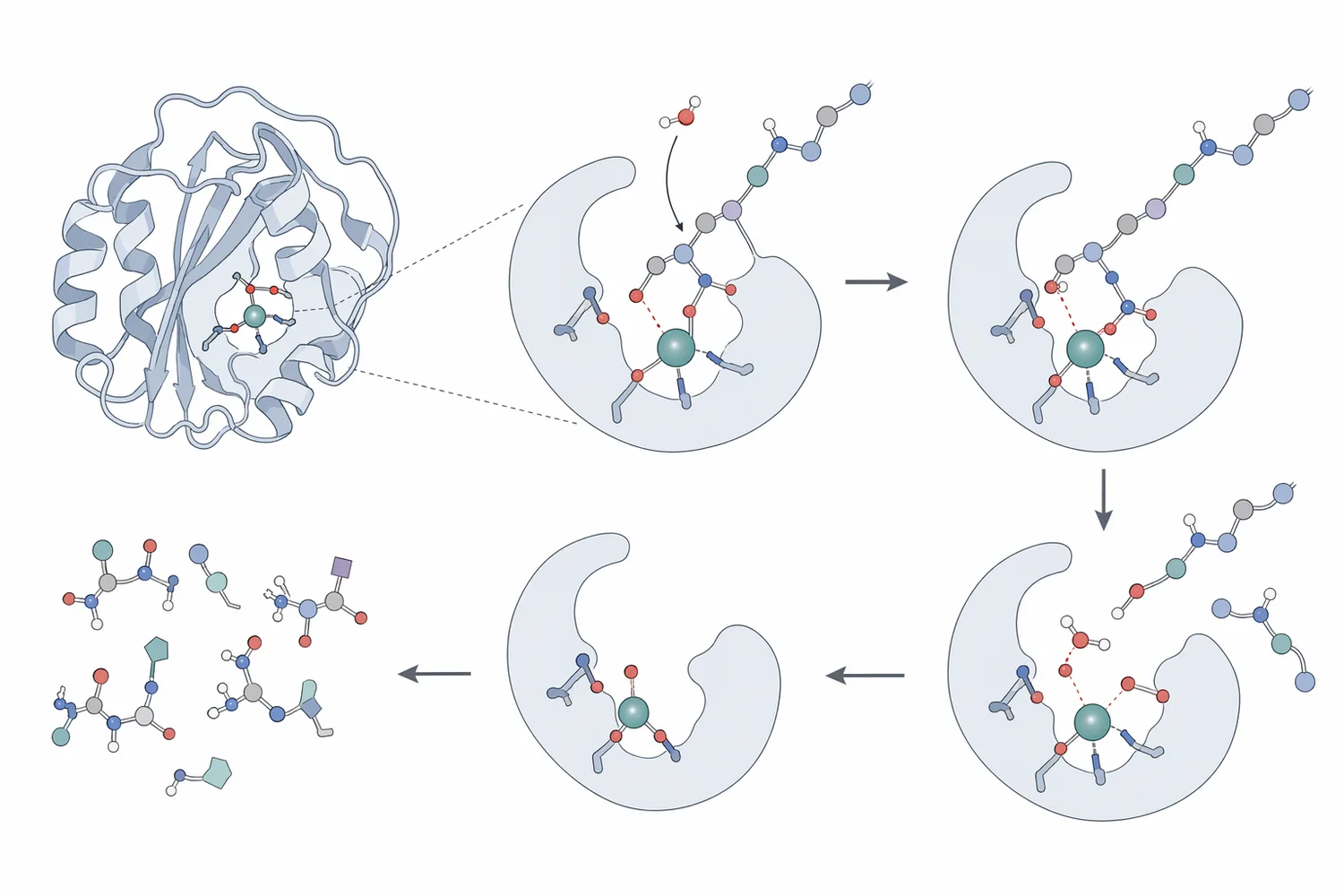
中性プロテアーゼは、タンパク質鎖中のペプチド結合を水の関与で切断します。大きなタンパク質は、折りたたみ構造、ジスルフィド結合、疎水性相互作用、塩橋、水素結合、他成分との複合体形成によって、水に分散しにくい状態を取りやすくなります。プロテアーゼが内部配列を切ると、分子サイズが小さくなり、凝集に寄与していた構造が崩れ、表面に露出する官能基の割合が変わります。その結果、溶解性、乳化界面への吸着、泡膜形成、粘度、消化酵素へのアクセス性、発酵微生物が利用できる窒素源量が変化します。豆類タンパク質の研究では、酵素加水分解がタンパク質構造を変え、機能特性の改善に結び付くことが整理されています[3]。
プロテアーゼの切断位置は無作為ではありません。各酵素には、切断部位周辺のアミノ酸配列、立体構造、局所的な電荷、疎水性、基質の折りたたみ状態に対する選好性があります。中性プロテアーゼの基質特異性を扱った研究では、酵素がどの配列を切りやすいかは活性中心と基質の相互作用に左右されることが示されています[4]。また、酵素の微小環境が変わると選択性や反応挙動が変化することも報告されており、同じタンパク質原料でもpH、塩、温度、共存成分、タンパク質の変性状態によって得られるペプチド分布は変わります[5]。
中性プロテアーゼの中には、金属イオンが触媒や構造安定化に関与するものがあります。たとえば、真菌由来中性プロテアーゼの生化学的特性を調べた研究では、特定の金属イオンや阻害因子に対する応答を通じて酵素の性質が評価されています[1]。ただし、「中性プロテアーゼ」という名称だけで、すべてが金属プロテアーゼである、あるいは同一の阻害剤感受性を持つ、と断定することはできません。食品・発酵・飼料用途では、名称よりも、対象原料に対してどの程度の分解パターンを生むかが工程品質を左右します。
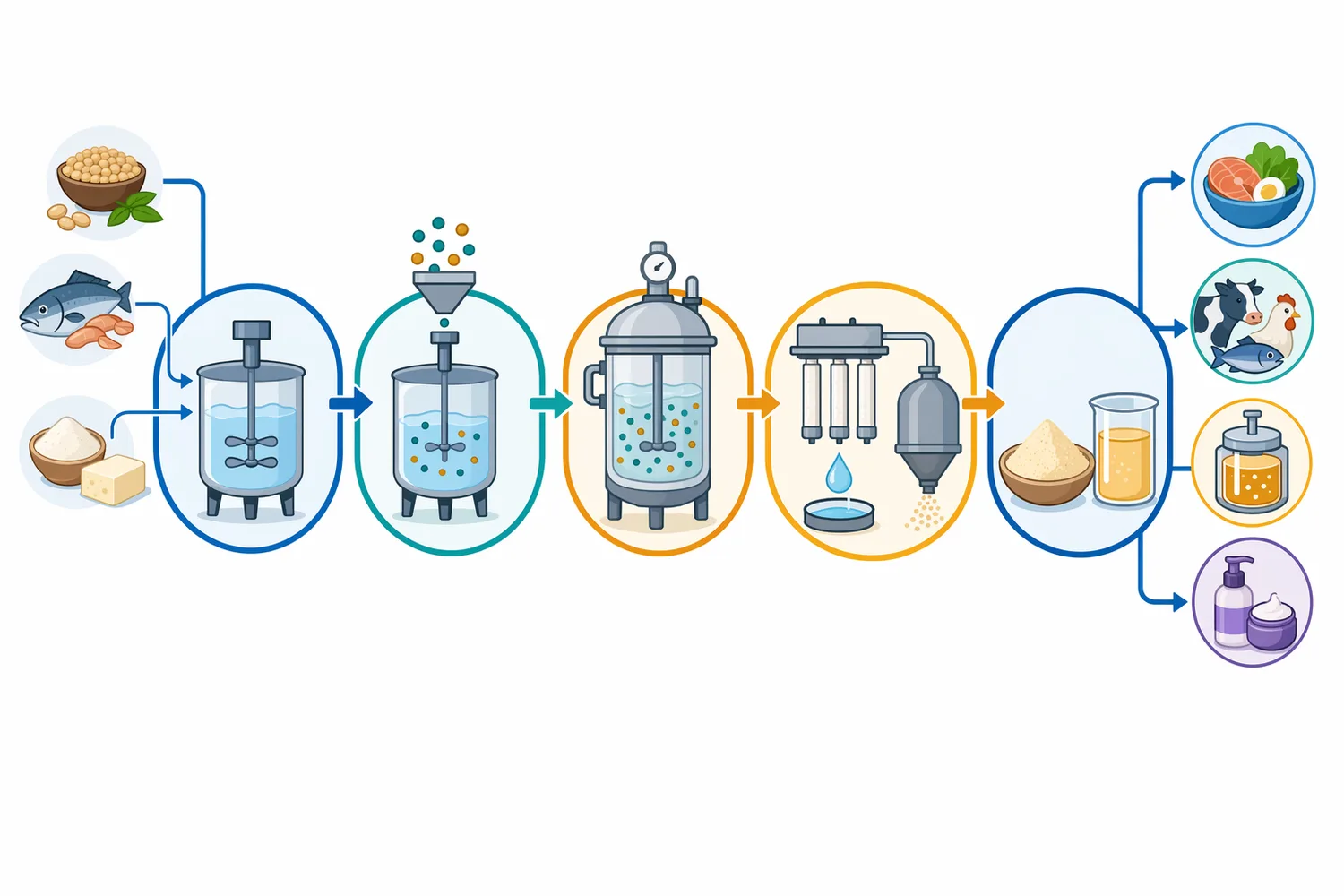
タンパク質加水分解で最も見えやすい変化は、溶解性と分散性です。高分子タンパク質が短いペプチドに分かれると、沈殿や凝集が抑えられ、粉末飲料、スープ、液体調味料、発酵エキスで扱いやすくなる場合があります。特に豆類・穀類・油糧種子タンパク質では、等電点付近の沈殿、加熱凝集、砂状感、口腔内のざらつきが問題になりやすく、酵素加水分解は分散性改善の一手段になります。パルスタンパク質のレビューでは、酵素加水分解が溶解性、乳化、起泡などのテクノ機能性を変えるツールとして扱われています[3]。
呈味も大きく変わります。プロテアーゼ処理により遊離アミノ酸や短鎖ペプチドが増えると、うま味、コク、発酵香気の前駆体、メイラード反応前駆体が増加することがあります。一方で、疎水性アミノ酸を多く含むペプチドは苦味に寄与しやすく、過度な加水分解は風味設計上のリスクになります。抗酸化ペプチド生産に関するレビューでも、酵素種、加水分解条件、原料タンパク質の違いが生成ペプチドの組成と機能性に影響することが整理されています[6]。
栄養・消化性の観点では、加水分解により消化酵素がアクセスしやすい短鎖ペプチドが増え、消化負荷の低減や吸収速度の変化につながる場合があります。乳タンパク質では、酵素加水分解と発酵が抗原性低減に使われることがレビューされており、エピトープを含むタンパク質構造の切断が抗原性変化に関与します[7]。ただし、アレルゲン性や免疫反応の低減は、特定のタンパク質、切断部位、残存エピトープ、製品マトリックスに依存するため、一般的な中性プロテアーゼ処理だけで一律に保証できる性質ではありません。
植物性タンパク質では、溶解性の低さ、豆臭、渋味、熱安定性、乳化・ゲル化のバランスが課題になります。中性プロテアーゼは、タンパク質を部分的に切断して分散性を上げたり、発酵前の窒素源を増やしたり、代替肉やプロテイン飲料の口当たりを調整したりする目的で利用されます。豆類タンパク質の酵素加水分解に関する研究では、加水分解によってタンパク質の構造が変化し、食品用途で重要な機能特性が改善される可能性が示されています[3]。
近年は、エンドウ、そら豆、レンズ豆、ひよこ豆、大豆などの植物タンパク質を、動物性タンパク質の代替素材として使う開発が進んでいます。これらの原料では、タンパク質の高分子構造と多糖・フェノール類・ミネラルとの相互作用が機能性を複雑にします。酵素加水分解は、タンパク質の分子量を下げるだけでなく、疎水性領域や荷電領域の露出を変え、乳化界面への吸着速度や水和性を変化させます。植物タンパク質加水分解のレビューでは、加水分解を通じて食品産業での用途拡大を図る考え方が整理されています[2]。
発酵工程では、微生物が利用できる窒素源の状態が増殖、酸生成、香気生成、ペプチド蓄積に影響します。中性プロテアーゼ処理でタンパク質を短いペプチドやアミノ酸へ変えると、発酵スターターが利用しやすい窒素源が増える場合があります。豆類タンパク質を乳酸菌発酵と組み合わせた研究では、微生物由来酵素による特異的加水分解、タンパク質組成、構造、機能性の変化が議論されています[8]。
発酵食品では、プロテアーゼ処理を単独で考えるより、微生物酵素、内在性酵素、塩分、水分活性、加熱履歴との組み合わせで見る必要があります。たとえば、発酵中に生成するペプチドは、うま味、苦味、甘味、酸味の感じ方に関与するだけでなく、メイラード反応や揮発性香気成分形成の前駆体にもなります。酵素処理によって分解を早められる場合がありますが、熟成香味はプロテアーゼだけで完結するものではなく、微生物代謝や後続反応が不可欠です。豆類発酵研究では、酵素分解と発酵がタンパク質構造および機能性を同時に変える複合プロセスとして扱われています[8]。
乳タンパク質では、消化性、抗原性、苦味、熱安定性、泡立ち、乳化、口当たりが設計対象になります。カゼインやホエイを中性プロテアーゼで部分加水分解すると、ペプチドサイズ分布が変わり、消化性や溶解性が変化します。乳タンパク質の抗原性低減に関するレビューでは、酵素加水分解と発酵が、抗原性タンパク質の構造を変化させる手段として整理されています[7]。
ただし、乳タンパク質加水分解では苦味管理が重要です。カゼイン由来の疎水性ペプチドは苦味を生じやすく、分解が不十分でも過剰でも好ましくない風味になることがあります。ホエイタンパク質加水分解物の研究では、加水分解条件と乾燥条件が加水分解物の特性や抗酸化性に影響することが示されており、酵素反応だけでなく後工程も最終品質を左右します[9]。
魚介残渣、内臓、皮、骨周辺組織、畜肉副産物は、タンパク質を多く含む一方で、臭気、脂質酸化、微生物負荷、加工適性のばらつきが課題になります。Neutral Proteaseによる加水分解は、これらの原料をペプチド、調味ベース、飼料素材、発酵基材へ変換する技術の一部として使われます。ティラピア副産物タンパク質加水分解物の研究では、酵素処理条件が抗酸化製品開発と品質設計に関わることが報告されています[10]。
魚介・畜肉由来の加水分解物では、うま味ペプチド、遊離アミノ酸、核酸関連成分、脂質酸化生成物が同時に風味へ影響します。プロテアーゼ処理はタンパク質部分を制御できますが、魚臭や酸化臭は脂質管理とも関係します。そのため、Neutral Proteaseは「副産物の価値化」を支える中心技術の一つであっても、最終製品の香味・色調・安定性は原料鮮度、脱脂、加熱、乾燥、発酵、ろ過などの工程と組み合わせて決まります。魚由来加水分解物の応用研究でも、タンパク質分解物の機能性評価は原料と工程条件に依存して扱われています[11]。
昆虫タンパク質や酵母タンパク質は、持続可能なタンパク質源として注目されていますが、消化性、色、風味、細胞壁構造、キチン、核酸、独特の香味などの課題があります。酵素加水分解は、これらの高タンパク原料をペプチド化し、飼料、食品素材、栄養素材として扱いやすくする技術です。食用昆虫タンパク質のレビューでは、酵素加水分解により生物活性ペプチドを生成し得る可能性が整理されています[12]。
酵母タンパク質加水分解物では、飼料添加物としての応用可能性が研究されています。酵母は細胞壁によりタンパク質へのアクセスが制限される場合があり、加水分解によって窒素成分の利用性や機能性が変化します。酵母タンパク質加水分解物の特性評価研究では、飼料添加用途を想定した栄養・機能面の検討が行われています[13]。
| 原料・用途領域 | 加工上の主な課題 | 中性プロテアーゼで狙う変化 | 注意すべき品質変化 |
|---|---|---|---|
| エンドウ・大豆・豆類タンパク質 | 溶解性不足、沈殿、豆臭、ざらつき、乳化不安定 | 部分加水分解による分散性、界面挙動、発酵利用性の調整 | 過分解による苦味、ゲル化力低下、粘度低下 |
| ホエイ・カゼイン | 抗原性、苦味、熱凝集、消化性設計 | ペプチド化による消化性・溶解性・抗原性関連構造の変化 | 疎水性ペプチド由来の苦味、後工程での反応性変化 |
| 魚介・畜肉副産物 | 臭気、脂質酸化、組織ばらつき、低利用価値 | ペプチド、調味ベース、発酵基材への変換 | 魚臭・酸化臭はタンパク質分解だけでは制御不可 |
| 発酵原料 | 微生物が使える窒素源不足、熟成時間、呈味不足 | ペプチド・アミノ酸供給による発酵促進や呈味前駆体形成 | 香味は微生物代謝と後続反応に依存 |
| 昆虫・酵母タンパク質 | 消化性、細胞壁、独特の香味、加工適性 | 高タンパク原料のペプチド化、機能性素材化 | 色・香味・不溶性成分の管理が必要 |
この比較から分かるように、Neutral Proteaseの価値は「同じ効果をすべての原料に与えること」ではなく、原料ごとの制約に対してタンパク質分子の大きさと構造を調整できる点にあります。豆類では溶解性や乳化性、乳では消化性や抗原性関連構造、魚介では副産物のペプチド化、発酵では窒素源供給が主な関心になります。植物タンパク質、乳タンパク質、魚介タンパク質、昆虫タンパク質のいずれでも、得られるペプチドの組成が機能性と官能品質を左右します[14]。
タンパク質加水分解物では、抗酸化性、ACE阻害性、抗炎症性、ミネラル結合性などの生物活性ペプチドが注目されています。中性プロテアーゼは、タンパク質鎖を切断して潜在的なペプチド配列を露出・放出するため、生物活性ペプチド探索の出発点として使われることがあります。豆類タンパク質加水分解物のレビューでは、抗酸化、血圧関連、代謝関連などの健康促進性が、生成ペプチドの配列と構造に依存して議論されています[14]。
ただし、生物活性は「加水分解すれば必ず得られる」ものではありません。酵素の切断特異性、原料タンパク質配列、処理条件、消化後の安定性、吸収性、食品マトリックス中での相互作用が関与します。コオロギタンパク質加水分解物から得られたテトラペプチドの研究では、抗酸化性とACE阻害性を示す配列が同定され、分子ドッキングも含めて作用が検討されています[15]。キヌアタンパク質加水分解物からの抗酸化ペプチド研究でも、精製・同定された配列の作用機序が個別に解析されています[16]。
ACE阻害ペプチドについても、原料ごとに異なる配列が得られます。スッポン肉水溶性タンパク質加水分解物から新規ACE阻害ペプチドを同定した研究では、特定配列が作用の中心として扱われています[17]。このような研究は、プロテアーゼ加水分解が機能性ペプチド開発の入口になることを示しますが、最終的な機能表示や生理作用の主張には、個別の配列同定、濃度、消化安定性、ヒトでの妥当性評価が必要です。
Neutral Proteaseを使う際に最も重要なのは、分解をどこで止めるかです。短い処理では高分子タンパク質が多く残り、溶解性や窒素源供給の改善が限定的になることがあります。処理を進めるとペプチドが増え、分散性や呈味が変わりますが、さらに進むと苦味、過度な低粘度化、機能性タンパク質の構造喪失が起こる可能性があります。ホエイタンパク質加水分解物の研究では、加水分解条件と乾燥条件の違いが最終特性や抗酸化性に影響することが示されています[9]。
もう一つの要点は、基質タンパク質の前処理状態です。加熱、せん断、pH調整、塩濃度、発酵、脱脂、粉砕によってタンパク質の折りたたみや凝集状態が変わると、プロテアーゼがアクセスできる切断部位も変わります。未変性の球状タンパク質では切断部位が内部に隠れていることがありますが、加熱変性やせん断で構造が開くと分解が進みやすくなることがあります。酵素特異性は酵素そのものだけでなく、基質の溶媒環境や立体状態にも影響されるため、同じNeutral Proteaseでも原料と工程によって結果は異なります[5]。
共存成分の影響も無視できません。ポリフェノール、多糖、脂質、塩、ミネラル、キレート性成分、還元剤、酸化生成物は、タンパク質構造や酵素活性に影響する可能性があります。特に植物タンパク質では、フェノール化合物や食物繊維との相互作用がタンパク質の可溶化や酵素アクセスを制限することがあります。未熟バナナ粉とデンプンの酵素加水分解抵抗性に関するレビューはタンパク質ではなく炭水化物系の例ですが、食品マトリックス中の構造、相互作用、物理的アクセス性が酵素反応を左右するという点では共通した示唆を与えます[18]。
近年のタンパク質加工では、酵素加水分解を単独で使うだけでなく、発酵、加熱、超音波、高圧、押出、膜分離、乾燥などと組み合わせる設計が増えています。抗酸化ペプチド生産に関するレビューでは、新規加工技術と酵素加水分解を組み合わせることで、タンパク質構造を変え、ペプチド生成効率や機能性を高める考え方が整理されています[6]。
たとえば、発酵とプロテアーゼ処理を組み合わせると、外部酵素による初期分解と微生物由来酵素による後続分解が重なり、遊離アミノ酸や小ペプチドの分布が変化します。乳酸菌発酵豆類タンパク質では、微生物の酵素系がタンパク質組成と構造を変え、機能特性に影響することが報告されています[8]。一方で、加水分解後の乾燥はペプチドの凝集、溶解性、酸化、褐変、抗酸化性に影響するため、粉末化を伴う製品では反応工程と乾燥工程を一体で考える必要があります[9]。
中性プロテアーゼ処理で最も典型的なリスクは苦味です。苦味は、疎水性アミノ酸を多く含む短鎖ペプチド、特定の末端配列、原料由来のフェノール化合物、脂質酸化生成物などが複合して生じます。プロテアーゼはペプチドを増やすため、うま味やコクを高める一方で、望ましくない苦味も増やす可能性があります。抗酸化ペプチドのレビューでも、酵素種と条件によって生成ペプチドの配列と特性が変わることが示されています[6]。
過分解は、機能性タンパク質素材では特に注意が必要です。乳化や起泡には、界面に吸着し、膜を形成できる一定の分子サイズと柔軟性が必要です。分解が浅すぎると溶解性が不足し、深すぎると界面膜を支える分子長が不足する場合があります。植物タンパク質の機能性改善では、加水分解により溶解性が上がる一方、乳化安定性やゲル化性は分解度に応じて増減するため、用途ごとの最適域が存在します[2]。
原料差も大きな要因です。同じ「大豆タンパク質」でも、分離タンパク、濃縮タンパク、脱脂粉、発酵原料、加熱履歴のある副産物では、タンパク質構造と共存成分が異なります。魚介副産物では部位、季節、鮮度、脂質量、結合組織量が異なります。昆虫タンパク質では種、飼育条件、脱脂、キチン含量が影響します。食用昆虫タンパク質のレビューでも、原料と加水分解条件が得られる生物活性ペプチドの種類に影響することが示されています[12]。
Enzymes.bioは、Protein Hydrolysate Enzyme Neutral Protease(CAS 9040-76-0)を1kg単位でオンライン直接販売する酵素供給業者です。製造業者や研究所としてではなく、B2B向けに酵素を購入しやすい形で提供する立場です。オンライン注文後に出荷処理が行われ、CoAおよびSDSは注文時に併せて提供されます。
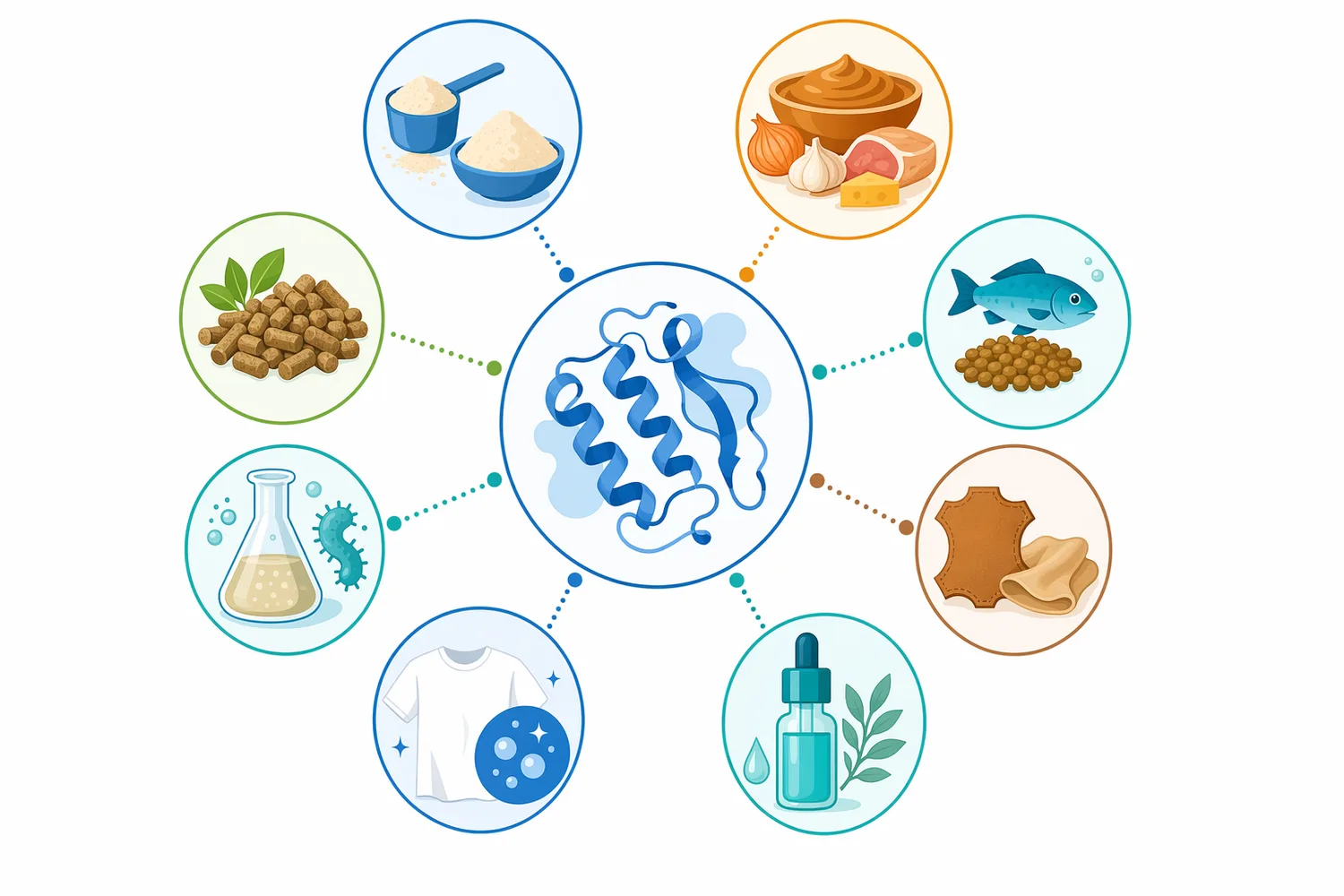
本製品は、タンパク質加水分解物、植物性プロテイン素材、発酵原料、調味ベース、乳・魚介・畜肉・酵母・昆虫タンパク質のペプチド化など、中性付近でタンパク質分解を行いたい工程に適した酵素カテゴリです。具体的な成果は、原料、工程、目的品質、後処理によって変わりますが、公開研究は、酵素加水分解がタンパク質の構造、機能性、呈味、消化性、生物活性ペプチド生成に大きく関与することを一貫して示しています[2]。
Protein Hydrolysate Enzyme Neutral Protease(CAS 9040-76-0)は、中性付近でタンパク質のペプチド結合を切断し、ペプチドやアミノ酸を生成するタンパク質加水分解向け酵素です。植物性タンパク質では溶解性・分散性・発酵適性、乳タンパク質では消化性や抗原性関連構造、魚介・畜肉では副産物の価値化、昆虫・酵母では新規タンパク質素材の利用性向上に関わります。酵素加水分解が食品タンパク質の機能性と用途拡大に寄与することは、植物タンパク質、豆類、乳、昆虫、魚介など複数の領域で研究されています[3][14]。
一方で、Neutral Proteaseは一律の品質改善剤ではありません。分解の深さ、切断選択性、原料の前処理状態、共存成分、発酵や乾燥などの後工程によって、溶解性、乳化性、苦味、香味、消化性、生物活性ペプチド生成は大きく変わります。したがって、本製品は「タンパク質を目的に応じた分子サイズとペプチド分布へ調整するための加工酵素」として使うのが最も適切です。Enzymes.bioでは、1kg単位のオンライン購入に対応し、注文時にCoAおよびSDSが提供されます。
1 kg単位で販売、在庫あり・即出荷可能です。オンラインストアで直接ご注文・決済いただければ、当社でご注文を処理します。すべてのご注文に試験成績書(CoA)と安全データシート(SDS)が付属します。
Protein Hydrolysate Enzyme - Neutral Protease 100000U/G Cas 9040-76-0を購入 →初出引用順に番号を付けています。各出典はオープンアクセスで、公開時にアクセス可能であることを確認済みです。本文中の引用番号からこちらにリンクしています。